He says "You missed the point, Mickey. What I am asking you to do is adjust my tree topology to suit your hypothesis. Then we can compare your changes to the large reptile tree. I’ll put up your figures and compare them with mine. Then we’ll look at synapomorphies. The list of characters is infinite for any taxon, as you can get down to individual chemistry and DNA if you like. So, merely listing a few to a dozen characters is not a solution. Rearrange the tree topology like you want to, then we’ll talk." And "You have your assignment: Nest Daemonosaurus with theropods while including Heterodontosaurus and Massospondylus. That’s a half-dozen to a dozen taxa at most to deal with. Then we’ll compare answers."
So that's what we're doing today. Taking the dinosaur portion of his tree, adjusting the codings to be correct and the character states to be well-formed, less correlated, ordered when appropriate, and seeing the results. Then looking at what synapomorphies support clades in Peters' tree.
For those of you who don't want to wade through the minutia below, here's the short answer. Peters'dinosaurian topology is completely due to miscodings and lack of characters/taxa. When coded right, poposaurs fall outside Dinosauria, and Trialestes and SMNS go next to Terrestrisuchus once it's added, but we get new odd things like poposaurian Daemonosaurus, and theropodan Pseudolagosuchus, Silesaurus and Saturnalia. As we add taxa one at a time, the phylogeny changes constantly so that sauropodomorphs are variously paraphyletic to theropods and/or ornithischians in different configurations, Saurischia is para- or monophyletic, silesaurs are polyphyletic in different ways, etc.. Simply put, there aren't enough characters to to stabilize topology, and it's only stable in Peters' matrix because he miscodes a ton. If we add the saurischian and dinosaurian characters I noted long ago on this blog, the topology is fairly standard, and if we add the character evidence for Daemonosaurus being a theropod, it becomes a theropod. I have no reason to believe the rest of his matrix isn't miscoded to a similar extent in similar ways, and that even if it were corrected, would lack the characters and taxa needed to be useful.
Methods
This evaluation uses Peters' amniote matrix from January 22, 2013. He has since updated it (I assume in part due to my comments on his site about miscodings in some of his 'theropod' OTUs), but the only difference is that Trialestes is now sister to Dinosauria instead of the most basal theropod. His matrix includes 228 characters, and this evaluation uses all of the taxa Peters recovers as avemetatarsalians (closer to birds than crocs). For outgroups, I chose Gracilisuchus and Turfanosuchus. Gracilisuchus is one of the most basal pseudosuchians (closer to crocs than birds) in Peters' tree, while Turfanosuchus is sister to Pseudosuchia+Avemetatarsalia. Both are also well described and known from articulated skulls. Note this is favoring Peters, as better outgroups in traditional phylogenies like Nesbitt's (2011) would be lagerpetids, pterosaurs and ornithosuchids. But Peters finds those taxa to be more distantly related, so they are ignored here. Similarly, Peters recovers Saltopus and Scleromochlus as pseudosuchians, so they are ignored. Another important point is that he finds Lewisuchus to be outside Archosauria, so it is ignored here though traditionally it is considered close to and probably synonymous with the included taxon Pseudolagosuchus. If we run this reduced analysis of Avemetatarsalia plus two outgroups, we get the same result as running the entire analysis of 328 taxa, so we know any changes in topology aren't due to only analyzing this area of the tree.
Running Peters' data results in one MPT of 354 steps. 90 characters don't vary in included taxa, and 18 variable characters can't affect the topology, so the number of useful characters is 120. It has a consistency index of .52, so on average each character reverses or converges once.

- For Turfanosuchus, only the type species was coded. The more fragmentary T. shageduensis is no longer thought to be congeneric. I lack the original description of Turfanosuchus, so this is the single instance where my codings do not reflect the total published knowledge of a taxon, but Peters lacks it too, so this shouldn't matter for our test. Nesbitt says it's not great anyway, and as it's in Chinese only the figures would be useful to me if I had it.
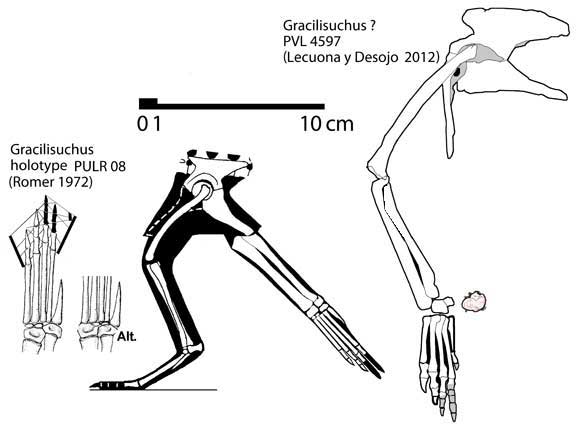
- For Gracilisuchus, I included all specimens generally attributed to it (the holotype PULR 08, MCZ 4116, MCZ 4117, MCZ 4118, PVL 4597 which is the Tucumen specimen, and PVL 4612). Romer (1972) used all of these when describing and illustrating the taxon, Brinkman (1981) reinterpreted the skull based on MCZ 4116 and 4118, and Lecuona and Desojo (2012) described the pelvis and hindlimb of PVL 4597 in depth. Peters did not realize Romer used these latter three specimens too, and coded PVL 4597's pelvis and hindlimb and most recently MCZ 4116+4118's skulls and cervicals as separate OTUs from Romer's composite description/drawings.
- For Trialestes, I only included the holotype, as Ezcurra et al. (2008) found the referred specimens to be a mix of dinosaur and indeterminate suchian bones (PVL 2559) and an indeterminate crocodylomorph (PVL 3889). This is important because Peters' codings are almost entirely the crocodylomorph holotype skull and the dinosaurian and possibly herrerasaurid pes of PVL 2559. So it's not surprising it falls out between crocodylomorphs and dinosaurs in his tree, since it's a composite.
- SMNS 12352 includes both a skull and a manus (these are the specimens originally referred to Procompsognathus that Sereno and Wild referred to Saltoposuchus in 1992), though Peters ignores the latter.
- Coelophysis is only coded based on C. bauri, which I think is what Peters did too.
- Massospondylus k is M. kaalae, shortened due to PAUP's display. This species is based on a partial skull.
- On Peters' Thecodontosaurus page, he illustrates both the type species T. antiquus and what was originally named T. caducus, which is now known as Pantydraco caducus. T. antiquus is based on a large number of generally disarticulated and unassociated bones, with a dentary as the holotype. Most recently, Galton (2007) redescribed them and found them to belong to at least three different taxa based on humeral morphologies. He named Asylosaurus based on the one articulated partial skeleton there, retained Thecodontosaurus for the dentaries and one kind of humerus, and kept most of the material unassigned. The codings for Thecodontosaurus in Yates' matrices are apparently based mostly on a bonebed that remains undescribed, though he and Benton are working on it. Given this mess, I've coded "Thecodontosaurus" based solely on Pantydraco and refer to it with the latter name. It includes the only described skull and pes, and Peters' clearly used it when coding his OTU. Later I add the chimaerical "Thecodontosaurus antiquus" as a separate OTU as a test.
- I included Stormbergia in the Lesothosaurus OTU, as I agree with Knoll et al. (2009) that their differences are ontogenetic.
Many of Peters' characters are composites, in that they code for multiple variables. "Serrations large and tooth roots constricted" as a state, for example. These had to be split into multiple characters to remain valid, which led to ten "new" characters being made. This brings the total number to 238. It would need to be much higher if the inappropriate states describing the anatomy of non-dinosaurs were examined as well.
Characters that have states which follow an objective sequence (e.g. "2 sacrals" "4 sacrals" "3 sacrals", or "humerus longer than femur" "humerus subequal to femur" "humerus shorter than femur") were set to be ordered in PAUP. Thus, the program knows that e.g. a short humerus is more similar to a subequal humerus than it is to a long humerus. It's implicit in many of Peters' states such as "more than 4 premaxillary teeth", since under unordered assumptions, 5 teeth is just as similar to 2 teeth as it to 6 teeth. Ordering characters sometimes meant I had to rearrange the states, so state 2 in my matrix will not always be the same as state 2 in Peters' matrix.
Many of Peters' characters are correlated with each other. Often this is due to one character coding for the presence/absence of a feature, then another character coding for attributes of that feature and also including a state coding for its presence/absence. This weights the presence/absence compared to other characters, and is easily solved by making taxa without the feature coded inapplicable for characters about that feature. Unfortunately, several correlated characters were retained, as altering them would lead to effectively using different characters than Peters, so it would be less fair of a test. For example, there's a character comparing each cranial fenestra size to orbit size. So if two taxa are exactly the same except one has larger orbits, they could be coded differently for "orbit compared to naris", "orbit compared to aof", "orbit compared to stf", "orbit compared to itf", and probably "orbit enters anterior half of skull" too. Similarly, there are several characters coding for the length of metatarsal V, separate characters for which digit is longest and which digit+metapodial is longest, and other such things. By not changing these, I'm giving Peters another advantage.
Results-
When those changes are made, we get one MPT of 458 steps. 95 characters don't vary in included taxa, and 31 variable characters can't affect the topology, so the number of useful characters is 122. It has a consistency index of .39, so has more homoplasy than Peters' codings. Differences include Daemonosaurus being a poposaur, poposaurs being outside Dinosauria, Silesaurus, Pseudolagosuchus and Saturnalia being theropods, Pantydraco being by Panphagia and Pampadromaeus, and Pisanosaurus being an ornithischian. But like Peters' original tree, Marasuchus, Trialestes and SMNS 12352 are still theropods, and Phytodinosauria still exists with some sauropodomorphs basal in it.

Let's try substituting the probably chimaerical Thecodontosaurus (including Thecodontosaurus, Asylosaurus at at least one other sauropodomorph as well) for Pantydraco. We get 95 MPTs of 453 steps. Thecodontosaurus clades with Massospondylus (due to massopod elements?), Herrerasaurus with Saturnalia, and there's much less resolution overall.

Adding taxa
As Thecodontosaurus is problematic, let's switch back to Pantydraco for the rest of the tests. Since Trialestes and SMNS 12352 are generally thought to be basal crocodylomorphs, let's add well known crocodylomorph Terrestrisuchus. It's thought by some to be a juvenile Saltoposuchus (which is poorly described), which is in turn thought by Sereno and Wild to be what SMNS 12352 belongs to. That gives us 180 MPTs of 490 steps.















Adding characters
Before we finish this, recall one of my posts criticized Peters analysis for leaving out 16 unambiguous synapomorphies of Saurischia and 11 such synapomorphies of Dinosauria (from Nesbitt, 2011). We've already seen that depending on the taxa you include, Peters' characters already can support a Saurischia containing all to most sauropodomorphs, but let's see what happens when we add those 16 saurischian synapomorphies. For those curious, I checked Nesbitt's accuracy too and found only 8% of the entries were miscoded, which is a fourth of Peters' total.



Comparing support for clades
Assignment completed, David. Let's compare answers. We'll compare the number of extra steps needed to find various groups in Peters' unaltered matrix, Peters' characters fixed and corrected by me using only the taxa he did, the fixed/corrected characters plus the 45 I added with all taxa added, and in Nesbitt's (2011; reduced to have only the taxa examined here that are shared) matrix. Note all of the constraint trees only involve taxa Peters' included, so e.g. the Dinosauria constraint doesn't force Sacisaurus out of the clade, though it does force Silesaurus out.
First, we'll enforce Avemetatarsalia/Dinosauromorpha, to the exclusion of traditional pseudosuchians.
Peters original- 30
Peters fixed- 2
Peters with new added- 0
Nesbitt- 0
Result- Peters didn't recover this clade based almost entirely on miscodings.
Next, enforcing Silesauridae+Dinosauria.
Peters original- 41
Peters fixed- 10
Peters with new added- 4
Nesbitt- 0
Result- Peters didn't recover this clade mostly due to miscodings, but also due to lacking certain characters and taxa.
Next, Dinosauria itself.
Peters original- 30
Peters fixed- 8
Peters with new added- 0
Nesbitt- 0
Result- Peters not finding traditional Dinosauria is due mostly to miscodings, but partially due to missing character and taxon data.
Peters' Phytodinosauria including Daemonosaurus, silesaurids and poposaurs.
Peters original- 0
Peters fixed- 12
Peters with new added- 34
Nesbitt- 76 (not Daemonosaurus)
Result- Peters' support is entirely due to miscodings, as fixing these makes the clade rather improbable. Adding characters and taxa makes this is extremely unlikely to be a real clade. And lest Peters claims for the nth time it's our emphasis on the crurotarsal ankle that blinds us to placing poposaurs in Dinosauria, even when proximal tarsal characters are excluded, this still takes 59 more steps in Nesbitt's analysis.
Saurischia.
Peters original-34
Peters fixed- 15
Peters with new added- 0
Nesbitt- 0
Result- Peters doesn't recover this due to half miscoding and half lack of included data.
Peters' Theropoda with Marasuchus and SMNS 12352.
Peters original- 0
Peters fixed- 2
Peters with new added- 19
Nesbitt- 34 (only Marasuchus)
Result- Peters recovered these as theropods due to miscodings, though adding taxa and characters makes them highly unlikely to be correct.
Peters' derived phytodinosaurs excluding Panphagia and Pampadromaeus.
Peters original- 0
Peters fixed- 16
Peters with new added- 35
*Cabreira et al.- 31
Cabreira et al.'s (2011) sauropodomorph analysis was used, as Nesbitt did not include Panphagia or Pampadromaeus. All taxa were retained, as none are outside the scope of this analysis except the outgroup Euparkeria.
Result- Peters recovered this completely due to miscoding. At 31-35 steps once other taxa and characters are added, it's highly unlikely to be correct.
'Paraornithischia' consisting of silesaurs, poposaurs and Pisanosaurus.
Peters original- 0
Peters fixed- 13
Peters with new added- 26
Nesbitt- 50
Result- Again recovered entirely due to miscodings, this becomes rather unlikely once recoded and extremely unlikely with added taxa and characters.
Daemonosaurus sister to Ornithischia sensu lato (Pisanosaurus ignored).
Peters original- 0
Peters fixed- 9
Peters with new added- 10
*Sues et al.- 7
Sues et al.'s (2011) analysis was used, as Nesbitt's does not include Daemonosaurus. Taxa outside the scope of this critique were deleted.
Result- Another grouping based entirely on miscodings, though in this case added taxa and characters don't affect its liklihood much. It's always moderately rejected.
Daemonosaurus in derived Theropoda (by Tawa and Coelophysis).
Peters original-18
Peters fixed- 5
Peters with new added- 0
*Sues et al.- 0
Result- Peters' analysis rejected this mostly due to miscodings, but once correct it's only weakly rejected. Adding taxa and characters makes it most parsimonious.
Sauropodomorpha+Ornithischia.
Peters original-14
Peters fixed- 2
Peters with new added- 8
Nesbitt- 13
Result- While Peters does not actually recover a traditional Phytodinosauria due to having Pisanosaurus, Pampadromaeus and Panphagia in the clade only if silesaurids and poposaurs are, the reduced version seems quite possible once his codings are fixed. Yet once more taxa and characters are added it becomes unlikely. In case you're wondering, making silesaurids phytodinosaurs is 28 steps less likely in both Nesbitt's matrix and the version of Peters with taxa and characters added.
Derived Sauropodomorpha+Ornithischia (Pisanosaurus ignored).
Peters original- 0
Peters fixed- 5
Peters with new added- 11
*Cabreira et al.- 25
Result- Peters has Saturnalia, Pantydraco/Thecodontosaurus and Massospondylus kaalae closer to ornithischians than Panphagia and Pampadromaeus. This is due to miscodings, though it still seems possible once these are corrected. More taxa and saurischian characters help make this unlikely, but what really kills it are the sauropodomorph charcters and intermediate taxa present in a large analysis like Cabreira et al.'s.
Sauropodomorpha.
Peters original-13
Peters fixed- 12
Peters with new added- 3
Nesbitt- 0
Result- Somewhat uniquely, Peters' characters and taxa really do reject Sauropodomorpha sensu lato (containing Panphagia and Pampadromaeus), even after correction. Though in the corrected version this is due to Saturnalia and Pampadromaeus being theropods, and Panphagia being outside Eusaurischia in the corrected version. Adding more taxa and characters makes Sauropodomorpha sensu lato only slightly unlikely, though Nesbitt's character selection makes it most parsimonious. Though Peters' alternative to monophyly was rejected soundly above (25 more steps in Cabreira et al.'s matrix), you might wonder about "my" alternative. This is also soundly rejected in Cabreira et al.'s matrix, taking 24 more steps.
Peters' arrangement of Ornithischia (Pis(Scel((Het+Hex)(Agi(Scut+Les))))).
Peters original- 0
Peters fixed- 8
Peters with new added- 14
*Butler- 12
The updated version of Butler et al.'s (2007) ornithischian analysis (from Coria et al., 2013) was used. All taxa were retained, as none are outside the scope of this analysis except the outgroup Euparkeria.
Result- Peters' odd ornithischian topology with Scelidosaurus basal is due to miscodings. Adding more data only increases its unliklihood, to a level similar to that in the most exhaustive published ornithischian analysis.
Butler's arrangement of Ornithischia (Pis(Het(Les(Agi+Hex)(Scut+Scel)))))
Peters original-18
Peters fixed- 9
Peters with new added- 3
*Butler- 0
Result-The current standard topology of Ornithischia is strongly rejected by Peters' original matrix, but still somewhat rejected after coding it correctly. Adding more taxa makes it only weakly rejected, and adding the many ornithischians and applicable characters from Butler's analysis supports it.
Character support
Peters requested we look at synapomorphies, so let's check what characters support heterodox nodes in his unaltered analysis. Of course based on the above, we know these nodes are due almost entirely to miscodings, but just to make sure the thrashing is total and unambiguous, we'll go in for a closer look...
To be continued...





{kind=link}
{kind=link}