 |
| Avemetatarsalia in Peters' most recent cladogram, taken from his website. |
I've attacked Peters' analysis from a number of angles. Noting it lacks characters that support traditional clades, noting its characters are poorly formed, noting numerous miscodings, but Peters remains unconvinced by these small assaults on portions of the data. He says "You missed the point, Mickey. What I am asking you to do is adjust my tree topology to suit your hypothesis. Then we can compare your changes to the large reptile tree. I’ll put up your figures and compare them with mine. Then we’ll look at synapomorphies. The list of characters is infinite for any taxon, as you can get down to individual chemistry and DNA if you like. So, merely listing a few to a dozen characters is not a solution. Rearrange the tree topology like you want to, then we’ll talk." And "You have your assignment: Nest Daemonosaurus with theropods while including Heterodontosaurus and Massospondylus. That’s a half-dozen to a dozen taxa at most to deal with. Then we’ll compare answers."
So nothing less than a total review of his data will do, it seems. And that's what I've done. Recoded all of the taxa he includes in Dinosauria in addition to some outgroups, gone through the characters and ordered/divided them when appropriate, and determined what Peters' characters and taxa actually show. What I've found is worse than I expected, but before we get into the results, I have a number of observations as to why his analysis is so bad.
First, Peters' codings are often not based on the preserved material at all. I'm not even talking about his Digital Graphical Segregation, i.e. overzealous tracing. No, I mean things like coding much of Turfanosuchus' postcrania from the Wikipedia photo of the specimen. So all of the manual and pedal characters are coded, just like you can see the bones restored at his website. But if he would have read Nesbitt (2011), he would know "only the proximal portions of the metatarsals are preserved, whereas the manus and most of the pes are sculpted", and indeed you can see the color difference of proximal metatarsals II-IV and phalanges IV-2 and IV-3 in the photo. Similarly, he codes much of Lotosaurus from mounts, when parts of these are fake and e.g. the pubis is actually unknown. Then there's the hand of Scelidosaurus, based on the specimen privately owned by Sole. But this is fake as the actual fossil does not preserve the manus (or most of the pes, or the tip of the snout...). Even worse is when he codes characters from areas not preserved or physically reconstructed. Like Pisanosaurus (whose skull is known from a posterior maxillary fragment), being coded for premaxilla-maxilla articulation, snout depth, lacrimal-naris contact, lacrimal height, squamosal extent, skull roof fusion, vomeral and parasphenoid teeth, premaxillary tooth size, etc.. Even his own skull reconstruction doesn't show these as known. Examples like this are common.
{kind=link}
 |
| Most preserved skull elements of Pantydraco, from Galton and Kermack (2011). Note combining these into a skull will not necessarily get you the result of Yates (2003) in every detail. |
{kind=link}
Peters also seems to code a LOT from reconstructions, without actually checking to see if the character can be determined from the material itself. Marasuchus restored with gastralia? Guess it had them, despite the fact it doesn't preserve any. Another good example is the skull of Pantydraco, where Yates (2003) provided a nice reconstruction, but most elements are incomplete with uncertain articulations with each other, and preserved with only one surface exposed (see figure above). A lot of reconstructions contain a good amount of plausible guesswork, but Peters treats it all as hard data to code. This is quite ironic considering that his own reconstructions often vary so much from published ones.
Connected to this flaw is Peters' overzealousness in coding characters that are based on the presence of small and easily lost elements. Since Peters' tree goes back to Ichthyostega, there are many characters coding for the loss of all of the fishy skull roof bones (supratemporal, postfrontal, intertemporal, postparietal, tabular, etc.). But taxa known from partial and disarticulated skulls are coded as lacking these, like Panphagia. The other big area affected by this are the small pectoral elements. Only preserve a disarticulated scapula and coracoid? Guess I'll code you as lacking clavicles, an interclavicle, sternal plates and cleithrum... Now sure all archosaurs probably did lack a cleithrum and most of the skull elements noted above, but it's just sloppy to code them that way when it's actually unknown. And sometimes the elements really are present, contra Peters' coding. Like Coelophysis having clavicles, or Heterodontosaurus and Scelidosaurus having ossified sterna. I wonder how much this kind of coding is affecting his tree down in areas where taxa actually do commonly have these elements.
 |
| All known skull elements of Panphagia (from Martinez and Alcober, 2009). Oh yeah, it definitely lacked supratemporals, tabulars, intertemporals and a postparietal.... |
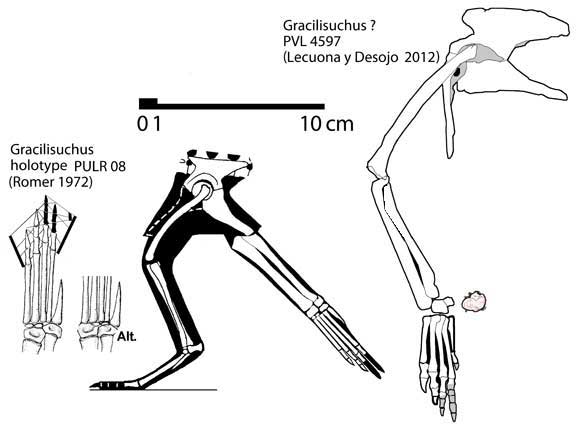 |
| Peters' comparison between supposed Gracilisuchus holotype (left) and Gracilisuchus specimen PVL 4597 (right). In actuality, the left is a composite between the holotype and PVL 4597, though the ilium and hindlimb of the holotype don't belong to Gracilisuchus. Note the "alternate" interpretation of Gracilisuchus as mesotarsal by Peters, which is a coded difference between the OTUs in his matrix. Not only is the illustrated tarsus based on (wait for it...) PVL 4597, but his alternate illustration flips the calcaneum horizontally! Sorry Peters, one side of that bone has a facet and the other doesn't, and it was quite clearly crurotarsal. Bones aren't just shapes that can be rearranged. But then, he jams the ischia in upside down against the pubes, though the latter were explicitly said to lack articular surfaces for the ischia. Sigh. |
In general, the coding suggests an unfamiliarity with the literature. For instance, Peters codes PVL 4597 (a specimen whose pelvis and hindlimb were recently described by Lecuona and Desojo) separately from Gracilisuchus as he believes it differs from the holotype. In the blog post, he compares the pelvis and hindlimb to figures 8 and 9 from Romer (1972), with the only noted difference being "Gracilisuchus has a metatarsal 4 shorter than 3. In the PVL specimen these two metatarsals were subequal." Yet that pes in Romer's paper is not the holotype, it's a composite between the holotype and what Romer called the Tucuman specimen. And what is the Tucuman specimen? None other than PVL 4597. Romer's text indicates the holotype doesn't preserve a complete metatarsal IV so the figure's IV/III ratio is based on PVL 4597. Peters was comparing the specimen to itself and finding it belonged elsewhere! The IV/III ratios of the left side as reported by Romer vs. L&D are 97% vs. 95%. A 2% difference, and the right side Romer hadn't prepared yet has a ratio of 105%. The other differences in his matrix are basically all due to miscodings.
This is just one example. There's the fact he doesn't code any skull characters for Poposaurus, when jaw elements are known. Sure, Parker and Nesbitt (2013) came out too late, but there's Gauthier et al.'s (2011) specimen which was coded into Nesbitt's matrix. Or that he codes Pisanosaurus for both cervical and caudal characters, when the vertebrae in question are a series that have been assigned to the neck or to the tail, but you can't have it both ways! Or that none of Hexinlusaurus' postcrania is coded, I can only assume because he was working with Barrett et al.'s (2005) brief discussion of the skull and lacked He and Cai's (1984) original osteology. Ditto for not having Padian and Rosenbaum's (2000) description of new Scutellosaurus material.
Yes it's hard to keep up with the literature, but it's necessary if you're going to proclaim sweeping heterodox conclusions as the truth.
As for the characters used, there are simply far too few, as I've always said. 228 sounds like a lot, but 96 of these only vary in taxa not examined here (e.g. cleithrum presence), and another 31 only vary in a single examined taxon, making them useless in this analysis (phylogenetically uninformative, to use the technical term). Which leaves us with 101 characters deciding dinosaur phylogeny. If we only use this same set of taxa in Nesbitt's (2011) analysis, it still has 199 informative characters. Even Cabreira et al.'s (2011) version of Yates' big sauropodomorph analysis (which has single OTUs for Crurotarsi and Ornithischia) has 159 informative characters, and Brusatte et al.'s (2010) archosaur analysis (which has two ornithischians and no sauropodomorphs shared with Peters') has 120. The characters that do vary are often basic proportions and shapes, with relatively few structural details (foramina, fossae, processes, laminae, etc.) coded. Thus it caters toward complete and articulated specimens, while more fragmentary taxa won't be able to be coded for much. For instance, Diodorus (known from partial dentary, teeth, humeri, femur and metatarsal III) can be coded for 34 characters in Nesbitt's analysis, but only 11 in Peters'. There's a huge emphasis on the skull (58% of the informative characters), and almost nothing on the main limb bones and tarsus. The latter could go a ways toward explaining why Peters doesn't recover pterosaurs close to dinosaurs. While usually we think of too few characters as being bad merely because taxa will be placed incorrectly without of the evidence being brought to bear, there are also more subtle consequences. For instance, if only relatively complete and articulated specimens preserve enough characters to code, then more fragmentary taxa that are important for being basal or having novel character combinations won't be able to be included.
Because most of the characters used are general proportions, they can more easily vary due to ontogeny and taphonomy. This is fine in larger analyses that will have more detail characters that can overcome them, but in Peters' analysis taxa often end up related due to comparative sizes of cranial fenestrae or limb proportions without regard to detailed anatomy. This probably explains in large part why Peters doesn't find standard clades in so many places.
 |
| What Peters' character 117 was intended to code. Mandibles in dorsal view of Captorhinus (left) and Proganochelys (right), showing surangular ridge in the latter highlighted in blue. Figure from Gaffney (1990). |
Several times Peters uses characters from standard amniote analyses, but misunderstands or misuses them. A few examples follow.
- Character 117 (Surangular lateral shelf: (0) absent; (1) present) refers to a laterally placed ridge in dorsal view- i.e. a narrow dorsal edge to the surangular, as described by Laurin and Reisz (1995- ch. 73). They used it to group procolophonids with turtles, and it can be seen clearly in e.g. Proganochelys (Gaffney, 1990- fig. 55, 56, 57B), while the lateral surface lacks a ridge (fig. 58, 59). Yet Peters codes it as if it refers to a ridge on the lateral surface that can be seen in lateral view, missing only in Pisanosaurus and poposaurs. In actuality, all scorable taxa have it, which is unsurprising as it is supposedly a eureptilian character as well.
- Character 45 is defined as "Frontal shape: (0) without a posterior process; (1) posterior process present. [deBraga & Rieppel 1997, Rieppel 1998]." Peters codes it as present basally but lacking in phytodinosaurs. Yet deBraga and Rieppel define the frontal lateral process as "a distinct lateral lappet (process) that projects out from the frontal margins at an abrupt 70° to 90° angle." This isn't present in any examined taxon.
- Peters defines character 50 as "Temporal ledge (distinct overhang at upper temporal area): (0) no ledge; (1) made from squamosal; (2) made from supratemporal and tabular. [Gauthier et al. 1988, Laurin & Reisz 1995, Hill 2005]." Yet none of those references include such a character. If he meant Gauthier et al.'s character 10 " Temporal musculature originates ventrally (0), or dorsally (1), on the parietal table" and Hill's character 91 "Temporal muscles on parietal table: (0) originate ventrolaterally – parietal margin straight; (1) originate dorsolaterally – parietal lateral margin embayed", that involves the medial edge of the supratemporal fenestra, not the lateral edge. The ledge is made from the parietal, not the squamosal, supratemporal or tabular.
While none of these actually vary in the examined taxa, I wonder what effect Peters' miscoding has deeper toward the tree base where they really do vary.
Another type of problematic coding involves classic characters Peters adds states to, which end up hiding homology. Imagine if we used the classic tetanurine character "less than X caudal vertebrae with transverse processes" but then added the state "caudals with elongated prezygapophyseal and chevron rods" and coded dromaeosaurids as having the rods. This would hide the fact they also share the low number of transverse processes with other tetanurines, so that it couldn't help group them together. A few examples from Peters' matrix follow.
- For character 187, taxa are coded either as having diverging pubic and ischial aprons (as opposed to a solid puboischiadic plate or a thyroid fenestra) or retroverted rods (in ornithischians). Yet ornithischians still have elongated pubic and ischial shafts instead of a solid plate or thyroid fenestra, regardless of whether the pubis is retroverted or missing an apron (both also covered by other characters anyway).
- Peters describes character 138 as swollen neural spines, and cites several authors. Yet those authors instead code swollen neural arches- Evans (1988- ch. B1), Laurin and Reisz (1995- ch. 86), deBraga and Rieppel (1997- ch. 106), Hill (2005- ch. 220), which means the width of the zygapophyseal portion of the neural arch. All of the taxa in this reduced matrix are coded a different state- transverse processes present. Yet the taxa the previous authors coded for their states have transverse processes too (e.g. Ophiacodon, Seymouria), so this distinction is invalid.
- For character 93, every scored taxon is coded as state 4- "large choana that virtually fill that portion of the palate." Oddly, the other states do not involve choana size. States 0 and 1 ("Choanae orientation: (0) parallel the maxilla; (1) posteriorly deflected medially") were taken from Laurin and Reisz (1995- ch. 40), deBraga and Rieppel (1997- ch. 8) and Hill (2005- ch. 113)- choana "(0) parallel to maxilla, palatine forms its posterior edge only; (1) curved posteromedially, palatine forms its posterior and part of its lateral edge." The scorable taxa actually have state 1 regardless of how big their choanae are.
As with the previous issue, this could really be affecting the phylogeny closer to the base of Peters' tree where the characters were designed to work.
 |
| Hypothetical example of two hands which would be coded the same by Peters' character 174 because in each a line intersecting the ends of metacarpals II and III intersects the end of manual phalanx I-1 as well. Lateral digits and phalanges on digits II and III left out for simplicity. |
The final kind of problematic character I'll outline here are those dealing with so-called interphalangeal lines. Peters (2000) published this idea, and it was well refuted by Hone et al. (2009). Quoting the latter authors, "interphalangeal (IP) lines connect sets of adjacent metatarsophalangeal (MP) or IP joints, and/or the tips of ungual phalanges. Peters (2000a) suggested that IP lines act as 'hinge lines' about which flexion and extension of the manus or pes can occur, that the lines might be helpful in matching fossil tracks to trackmakers and elucidating phylogenetic relationships, and that they might represent a powerful tool for vertebrate palaeontologists because the lines could be used to predict the lengths of missing phalanges." Even if Peters were right that interphalangeal lines could be objectively identified and were functionally useful, they would make terrible characters for phylogenetic analyses due to the multiple ways of achieving the same state. For example, in his analysis character 174 codes for where in manual digit I a line intersecting the ends of metacarpals II and III would go. But the angle of that line depends on the comparative lengths of metacarpals II and III (if II is longer the line angles distomedially, if III is longer it angles proximomedially), and where in digit I it intersects depends on the lengths of metacarpal I, phalanx I-1 and manual ungual I. Note the figure above. They would be coded the same, but that sameness isn't anything that could be accounted for by an inherited mutation. Hands and feet don't develop in a way that the lengths of metacarpals II and III could be connected to the lengths of I and phalanx I-1 at a certain angle. This goes to show not every objective feature should be used as a character in a phylogenetic analysis. An equivalent would be the hypothetical character "number of presacral vertebrae divided by number of sacral vertebrae, answer rounds to an - even number (0); odd number (1)." You'll get a definite answer, but the same state could be arrived at in many unrelated ways and is nothing a shared mutation could account for.
But hey, all of this is missing the point. What's the result of evaluating all of Peters' data for dinosaurs? Does it support his phylogeny? Stay tuned, as that's coming next time.
I'm on the edge of my seat...
ReplyDeleteMickey, I appreciate your thoroughness and your insight. It 'takes a village' to create a tree.
ReplyDeleteWith regard to Parallel Interphalangeal Lines (PILs), Hone et al. (2009) actually confirmed the presence of PILs several times throughout their paper, never tested it on a taxon with more than three toes, and demanded more precision in the digital lineups than nature can provide. Some taxa are actually known by their interrupted PILs. Others are in evolutionary transition from one pattern to another. And most of the time the PILs do not become apparent until they are used (as you'll see when you make a fist or grab a golf club, but not so much when you hold your hand flat). More on the foibles, mistakes and oversights of Hone et al. (2009) were published in Peters 2010, DOI: 10.1080/08912961003663500. (Funny how the rejoinders are never noted by you guys).
Your illustration of a possible problem in toe morphology provides keen insight, and would indeed score as a convergence in my matrix. It would not be the only convergence on the tree (the vast majority of traits used are convergent somewhere) and the suite of other characters would drown out that noise for maximum parsimony. (I sense you are using the old Larry Martin trick of "show me one character that unites birds and dinosaurs." It's always the suite of several hundred traits that provides the nesting site, not just a single or a dozen traits).
You found an interesting coding of the mandible shelf where I thought it was not. Is that shelf the same sort of shelf (homology) seen in higher taxa? If not, that's why I scored it differently.
I do score my matrix from reconstructions and mounts and therein lies a potential problem. If I'm not aware of some plaster, it gets scored as if real. In traits that are not real scores would tend to randomize, adding noise to the parsimony, not direction. I have often mentioned if better data is out there, please send it in. This goes double for plaster. If you have not done so, then please do so. I often find problems with my own coding and endeavor to update them. I will continue to do so as my tree evolves on its own. I am not married to the tree that I recovered, but it is the tree that I recovered and sister taxa do bear more than a passing resemblance to each other throughout the tree. Some branches are weaker. Others are quite strong.
Your earlier tree had a series of plant-eaters preceding theropods and separating theropods from bipedal, equally carnivorous crocs. I look forward to seeing what your new tree looks like and trust that you will have found a closer model to the actual evolutionary tree of archosaurs/dinosaurs. If Daemonosaurus doesn't lead to phytodinosaurs, I'd like to know the reason why and, if more parsimonious elsewhere, I'll buy you dinner. -- Dave
Thanks for the heads-up on the mistakes I made with the PVL specimen attributed to Gracilisuchus. I misread right for left on the ischium and believed Romer.
ReplyDeleteRegarding interphalangeal lines, I said it doesn't matter if these could be objectively identified (choosing the right angle to put each phalanx and metapodial from adjacent ones) or are functionally useful (as the roughly straight line between first phalanges on manual digits II-V is when we hold a golf club, as you say). My issue with them being used as characters is that morphological characters are basically stand-ins for mutations that could be inherited. Characters have to be heritable to make sense to use, and all we inherit from our parents' (in cultureless species like basal dinosaurs) is mutations. So what kind of mutation could code for metacarpals II and III lining up with phalanx I-1? We know how limbs form in archosaurs. Check Figure 4 of Wagner and Gauthier (1999). Digit IV forms, the next distal carpal splits off and III forms from that, then distal carpal II and digit II forms, then distal carpal I and digit I branches from that. For interphalangeal lines to be heritable, there would need to be a straight line in the embryo that told the mesenchyme when to disintegrate between future bones. What developmental mechanism do you know that works in straight lines between elements formed at different times?
ReplyDeleteSimilarly, the problem with the different hands in my figure isn't convergence. I expect some convergence in any tree. The problem is I fail to see how the two hands are similar in any way development or evolution could account for. Sure us humans can make a line from two bones and say it falls at a certain place in another bone, but just like with my vertebral equation character, I don't see how these two conditions are something nature could arrive at using the same mutations.
"Higher taxa" do indeed have that shelf, if you mean the archosaurs examined here. Turfanosuchus, Gracilisuchus, Shuvosaurus, Effigia, Herrerasaurus, Panphagia, Pisanosaurus and Heterodontosaurus all seem to have it, for instance. It could be convergent with Proganochelys for all I know, but that's the feature prior authors were coding for, not any shelf determinable from lateral view.
I'm glad you see scoring from reconstructions and mounts as a problem. With trees often differing from each other by only a few extra steps, we cannot afford to trust that wrong guesses will balance each other out. I include all references in my extensive character-by-character critique. If you lack one, ask me for it.
For Daemonosaurus, mostly miscodings of it from what I can tell so far. At least we're on the same coast, so dinner's not implausible. ;)
What developmental mechanism do you know that works in straight lines between elements formed at different times?
ReplyDeleteThe why and how questions never come up in phylogenetic analysis. IF the tibia was longer than the femur in a near legless taxon and in a cursor, that's a convergence. Wings, or a Carapace, as a trait, is a convergence. How they develop is not germane to scoring. It all comes out in the wash. The suite is what counts. And the choice of traits can skew the suite. That's one reason why I haven't added traits for the last 150 or so added taxa. I just want to see what happens with what I got.
They usually don't come up because authors are usually better at forming characters. Again I use my "number of presacral vertebrae divided by number of sacral vertebrae, answer rounds to an - even number (0); odd number (1)" fictional example. If this were in an analysis, you wouldn't say it doesn't matter how or why it's supposed to work, you'd say there's no way such a character could be genetically controlled.
ReplyDeleteYour philosophy toward character choice and coding in general is far too blase. You can't trust the phylogeny to "come out in the wash", certainly not with 228 characters. What you don't seem to realize is that (even when correctly coded), your choice of characters (and taxa) does skew the suite. If you "just want to see what happens" when using those characters, great, but then don't pretend it's giving you a well supported phylogeny.