As mentioned by Justin Tweet last month, we haven't had that many new dinosaur taxa described lately, whether due to COVID or random chance. But this doesn't bother me because there is always so much data to catch up on. In mid May I checked my Database suggestion emails and the most recent topic to address was "Brochu (2003) mentions that the specimen AMNH 6266 (including a jugal,
lacrimal, quadratojugal, and D-shaped premaxillary tooth) might belong
to the holotype of Alectrosaurus olseni as it was found at the type locality of A. olseni." And because of that, we now have all published Iren Dabasu theropod records added and updated on The Theropod Database. I'll go over some of the lesser known and new details about select taxa here. There was nothing particularly new about Erliansaurus, Neimongosaurus or Gigantoraptor, so I don't list them below. If anyone knows of any Iren Dabasu theropod specimens in the literature I missed, do leave a comment. One thing that struck me was just how much material remains undescribed, at the AMNH, IVPP and PIN especially. If anyone needs projects based on collected material- sort out Archaeornithomimus, describe the many therizinosaur elements at the IVPP, describe the partial troodontid skeletons at the AMNH, etc..
Alectrosaurus olseni
(see page for materials list)
The lectotype hindlimb AMNH 6554) was discovered on April 25 1923 at
Third Asiatic Expedition field site 136, while a partial forelimb (AMNH
6368) was found on May 4 at field site 138, 30 meters away. Andrews
(1932) first mention the former as "the complete hindlimb of a large
carnivorous dinosaur. The leg lay doubled up just as the great reptile
had died millions of years ago." Gilmore (1933) made each a syntype of
his new taxon of deinodontid, Alectrosaurus olseni,
noting "in the field they were thought to pertain to the same
individual." but that he preferred to treat them as two individuals.
They were considered the same taxon based on the manual unguals
(questionably associated in the case of AMNH 6554) "being laterally
compressed, strongly curved, and having sharply pointed extremities",
which are characters broadly true of almost all theropod manual
unguals. Barsbold (1976) was the first to consider AMNH 6368 wrongly
assigned, stating "As new materials from the MPR* show, a large ungual
phalanx previously attributed to the manus of Alectrosaurus
(Gilmore, 1933) does not really belong to it" (translated), citing the
still undescribed IGM 100/50 from Bayanshiree which includes "a small
ungual phalanx of the first manual digit, quite typical for
tyrannosaurids." Further, he noted "A large, laterally compressed
ungual phalanx, similar in structure and form to that attributed to Alectrosaurus,
belongs to another previously unknown dinosaur (under study) found
there. This dinosaur does not belong to Tyrannosauridae.", which is a
reference to the also undescribed Segnosaurus.
In their redescription, Mader and Bradley (1989) describe AMNH 6368 in
detail and place it in Segnosauridae, and it has been viewed as
therizinosaurian since then. As explained by Mader and Bradley, the
type listing by White (1973) combined the syntype materials, so that
Welles and Long (1974) officially declared the hindlimb as the
lectotype when they stated "we here designate this specimen, AMNH 6554,
the type of the species." Zanno (2010: Fig. 9D) figured the manual
unguals of AMNH 6554 as therizinosaurian without comment, although
their non-tyrannosaurid characters could also be plesiomorphically
shared with e.g. Dryptosaurus.
Dong et al. (1989) first reported Aublysodon
from the July 1988 Sino-Canadian expedition (CCDP), and Currie et al.
(1990) stated "Identical teeth [to Dinosaur Park juvenile tyrannosaurid
'Aublysodon'] recently were recovered from the Iren Dabasu Formation at Erenhot, People's Republic of China (IVPP 170788104). The Asian "Aublysodon" teeth belong to Alectrosaurus
(Perle pers. comm. 1989..." Similarly, Dong (1992) reports "In July
1988, the expedition of the CCDP came to Erenhot (Fig.85) where they
collected ... teeth of ... large theropods (tyrannosaurid)." Dong et
al. first reported that in July 1988 "A partial skeleton of Alectrosaurus was discovered too late to collect", and Dong (1993) followed that up by writing "An incomplete skeleton of Alectrosaurus
was found by Currie [in 1988], but was not excavated until the return
expedition of 1990", which was at CCDP site #9 based on Currie and
Eberth's (1993) table 3. They further noted "Perle (1977) ... has been
studying more recently discovered postcranial specimens (Perle, pers.
comm. 1989)" and that "The absence of denticles on the premaxillary
teeth (Perle, pers. comm. 1989; IVPP 180788-104) suggests that it
should be included in the Aublysodontinae." The similar field numbers
to Currie et al.'s suggest one is a typo, and serrationless
premaxillary teeth are a juvenile character of tyrannosaurines but also
known in some basal tyrannosauroids (Yutyrannus, Xiongguanlong). Currie (2001) reported "Several partial, undescribed skeletons of Alectrosaurus
collected from southeastern Mongolia are in the collections of the
museum in Ulaanbaatar, and another new specimen was recently collected
from Erenhot in China", the latter seemingly being the one mentioned by
Dong et al..
(see page for references)
undescribed tyrannosauroid (Granger and Berkey, 1922)
Middle-Late Campanian, Late Cretaceous
Iren Dabasu Formation, Inner Mongolia, China
Material-
(AMNH 6266; "AMNH 6556" of Carr, 2005) (small) incomplete lacrimal,
anterior jugal, quadratojugal, lateral ectopterygoid, posterior
pterygoid, premaxillary teeth, lateral teeth
Comments- Brochu (2003) noted "A box of bone fragments (AMNH 6266) from the same locality [as Alectrosaurus]
includes small tyrannosaurid skull bones (including a characteristic
jugal, lacrymal, quadratojugal, and D-shaped premaxillary tooth) that
might belong to the same individual. ... The skull parts are consistent
with Albertosaurus; for example, the jugal foramen is a dorsally-opening slit." He states it "had been originally catalogued as "Deinodon
sp." ..., but this was subsequently scratched off and "Theropoda
indet." written on in pencil", and the AMNH online catalogue does list
6266 as "Deinodon
? sp." Carr (2005) later reported "an undescribed, but shattered,
tyrannosauroid skull (AMNH 6556) from the same general area - Iren
Dabasu -" as the Alectrosaurus
lectotype, but as they are from different locations "there is no
evidence they are from the same individual." Carr states "The presence
of a secondary fossa in the antorbital fossa of the jugal indicates the
specimen is referable to Tyrannosauridae. The cornual process of the
lacrimal is similar to some juvenile tyrannosaurids in that it is a
low, laterally extending ridge. The lateral teeth are as finely
denticulate as tyrannosauroid teeth of the same basal crown length from
the Turonian of Uzbekistan." This is the "Iren Dabasu taxon" in Carr's
tyrannosauroid analyses as of Carr et al. (2017), under study by Carr
and recovered in a
polytomy with Timurlengia, Jinbeisaurus
and eutyrannosaurs as of 2019. While this is certainly the same
specimen based on material preserved, the AMNH online catalog lists
this specimen number as being a saurischian metatarsal II with a
locality "8 mi. E. of station" which would place it among Third Asiatic
Expedition field sites 140-149 (while Alectrosaurus
is from 136 less than a mile south of the station). AMNH 6556 is
listed in the catalogue as collected on April 30, which matches Carr's
statement the skull was found in late April five days apart from Alectrosaurus'
lectotype (which was found on April 25). However, Mehling (pers. comm.
6-2022) indicates AMNH 6556 is actually a metatarsal II and that AMNH
6266 was discovered in 1922, so that Carr apparently got the specimen
number wrong and incorrectly used the metatarsal's discovery date for
the skull. The early discovery makes sense considering the low
specimen number and allows us to equate the material with "portions of
a small carnivorous dinosaur skull with two or three teeth" found in
the 1922 expedition as reported by Granger and Berkey (1922) along with
ornithomimid remains that are near certainly AMNH 6267-6268. If it was
recovered with the latter specimens, AMNH 6266 would have been found
between April 25 and May 7 at one of the western AMNH quarries
(131-138), and thus may be from the same locality the Alectrosaurus
type as stated by Brochu. Alas, the only recorded locality information
in the AMNH card catalogue is Iren Dabasu (Mehling, pers. comm. 6-2022).
References-
Granger and Berkey, 1922. Discovery of Cretaceous and older Tertiary strata in Mongolia. American Museum Novitates. 42, 7 pp.
Brochu, 2003. Osteology of Tyrannosaurus rex: Insights from a nearly
complete skeleton and high-resolution computed tomographic analysis of the skull.
Society of Vertebrate Paleontology Memior. 7, 138 pp.
Carr, 2005. A reappraisal of tyrannosauroids from Iren Dabasu, Inner Mongolia,
People's Republic of China. Journal of Vertebrate Paleontology. 25(3), 42A.
Carr, Varricchio, Sedlmayr, Roberts and Moore, 2017. A new tyrannosaur
with evidence for anagenesis and crocodile-like facial sensory system.
Scientific Reports. 7:44942.
undescribed Tyrannosauridae (Gilmore, 1933)
Middle-Late Campanian, Late Cretaceous
Iren Dabasu Formation, Inner Mongolia, China
Material- (AMNH 21552) (large) femur (Mader and Bradley, 1989)
(AMNH coll.) (large) pedal elements (Gilmore 1933)
(IVPP coll.) teeth and/or elements (Yao, Wang, Sullivan, Wang, Stidham and Xu, 2015)
Comments- Under Deinodontidae, Gilmore (1933) states "The presence of a second carnivore, apparently rivaling Tyrannosaurus
in size, is indicated by a few scattered foot bones." These are no
doubt one or more of the specimens listed here under undescribed
Averostra (e.g. AMNH 6376, 6556, 6744, 6756, 6757, etc.).
Mader and Bradley (1989) noted "among the materials brought back by the
Central Asiatic Expeditions was the isolated femur (AMNH 21552) of a
much larger tyrannosaur [than the Alectrosaurus lectotype]." This may belong to the same taxon as Gilmore's pedal material based on size.
Yao et al. (2015) note "small unarticulated bones and teeth, including
fossils of ... tyrannosauroids" from "a rare microvertebrate locality
within the Iren Dabasu Formation, about 16 km northeast of Erenhot
City."
References- Gilmore, 1933. On the dinosaurian fauna of the Iren Dabasu
Formation. Bulletin American Museum of Natural History. 67, 23-78.
Mader and Bradley, 1989. A redescription and revised diagnosis of the syntypes
of the Mongolian tyrannosaur Alectrosaurus olseni. Journal of Vertebrate
Paleontology. 9(1), 1-55.
Yao, Wang, Sullivan, Wang, Stidham and Xu, 2015. Caenagnathasia
sp. (Theropoda: Oviraptorosauria) from the Iren Dabasu Formation (Upper Cretaceous:
Campanian) of Erenhot, Nei Mongol, China. Vertebrata PalAsiatica. 53(4), 291-298.
 |
| Iren Dabasu ornithomimosaur pubes (left to right) - AMNH 21799 figured by Smith and Galton (1990) as Archaeornithomimus asiaticus so generally associated with the taxon; AMNH 21798 and a distal pubis in AMNH 6570, both among Archaeornithomimus material from the same bonebed as 21799; LH-02-01 from an unknown locality in the same area described by Yao et al. (2022) as Ornithomimosauria indet.. The first three are my photos courtesy of the AMNH (scale = 100 mm), the fourth is after Yao et al., 2022. |
Archaeornithomimus asiaticus
(see page for materials list and additional comments)
Comments- Watanabe et al. (2015) state "Based on manual articulation of
disarticulated sacral vertebrae, we identify the two sacral vertebrae
(AMNH FARB 21790) sampled for this study as the second and third sacral
vertebrae", but these are the second and third caudals as identified by
Smith and Galton as confirmed by personal observation when sacrals 2-5
were on loan articulated to the ilium. Similarly, two of the proximal
caudals Watanabe et al. call AMNH 21790 are actually the last two
vertebrae in AMNH 21802.
Other material- The AMNH catalog lists 6267 and 6268 as Ornithomimus sp., but they are
probably Archaeornithomimus
based on stratigraphy. Given their lower specimen numbers, it is
likely these were recovered in the initial reconnaissance expedition to
Iren Dabasu in 1922 (April 25 to May 7) and represent the "Carnivorous
dinosaurs of at least two genera, the smaller one being of the Ornithomimus type" reported by Granger and Berkey (1922). Indeed, those authors later state "Remains of the small Ornithomimus-like
creature are particularly abundant and the last day at Iren Dabasu we
picked up probably fifty good foot bones and centra from two or three
knolls", which matches the listed material. The locality of AMNH 6267
is listed as "Iren Dabasu Sta. 1 1/2 SW of auto trail", so was
plausibly from one of the western AMNH quarries (131-138), as opposed
to the type material that was from the Kaisen Quarry (AMNH locality
140) and Johnson Quarry (AMNH locality 141) which were discovered the
following year. The AMNH online catalogue also lists AMNH 21626 and
21627 as possibly referrable to Archaeornithomimus. AMNH
21597 is figured as therizinosaurid ungual on the AMNH online catalog,
but the low curvature and distally placed flexor tubercle are instead
almost identical to Archaeornithomimus
(e.g. AMNH 6570, 6576). Currie and Eberth (1993) state "The present
whereabouts of a partial skull found by the Sino-Soviet expedition is
currently unknown", but their paper's details indicate it was found in
June 1959 from their localities K (= AMNH locality 141?), L or P, and
initially stored in either the IVPP or PIN. Chow and Rozhdestvensky
(1960) specified the timing of excavations to be June 14 to July 17 and
state "materials collected include ... small ornithopods (of Struthiomimus
type)", with 'ornithopods' presumably a typo for 'theropods', and
Currie and Eberth state "more than a thousand bones" were identified as
ornithomimid in the filed from the Sino-Soviet expedition. Currie and
Eberth also say that after the joint BMNH - Inner Mongolian Museum
expeditions of 1972-1977, "Some of the specimens (including
ornithimimid ... skeletons) were prepared for display in Hohhot", with
casts at the Erenhot Dinosaur Museum. These were mostly from
localities on the west side of Iren Nor. Dong (1992) noted that "Archaeornithomimus
is the most common species in the bone beds" where the CCDP excavated
in July 1988 (and later in 1990), which would have ended up in the IVPP
as no theropod body fossils from Iren Dabasu are at the TMP. A distal
caudal described by Makovicky (1995) as Avimimus "has the morphology of a typical
coelurosaurian distal caudal, but is otherwise undiagnostic. The
possibility that it may originate from an avimimid is suggested by its
small size. It should be noted, however, that it could just as
conceivably be from the tail of a juvenile Archaeornithomimus from the same bonebed." Indeed, the A. nemegtensis bonebed show Avimimus has short distal caudals like other caenagnathoids and unlike ornithomimosaurs, so this specimen is here referred to Archaeornithomimus. Godefroit et al. (1998) reported "one single ornithomimid caudal vertebra" from the Bactrosaurus
bonebed they described from Locality SBDE 95E5, slightly to the west of
AMNH locality 140. Yao et al. (2015) note "small unarticulated bones
and teeth, including
fossils of ... ornithomimids" from "a rare microvertebrate locality
within the Iren Dabasu Formation, about 16 km northeast of Erenhot
City."
More than one taxon? There is non-ornithomimid material catalogued under Archaeornithomimus,
including a juvenile ?Bactrosaurus ungual (in AMNH 6576),
a small ?troodontid pedal ungual I (in AMNH
6576) and part of an ?oviraptorid manual ungual I (in AMNH 6570).
Yao et al. (2022) described pelvis and sacrum LH-02-01 discovered in
2002 from an undocumented locality near Iren Nor. They added the
specimen to Choiniere's coelurosaur analysis to recover it as an
ornithomimosaur in a polytomy with Nqwebasaurus, Pelecanimimus, Shenzhousaurus, Beishanlong and Deinocheiridae+Ornithomimidae. Supposed differences from Archaeornithomimus
are- larger size (ilium 342 vs. 114 mm); "the shortest sacral centrum
is the second rather than the third [actually the third instead of the
fourth, assuming six sacrals as in Archaeornithomimus-
Makovicky, 1995]; the centrum of the first caudal is shorter than that
of the fifth [sixth] sacral"; first caudal with flat posterior
articular surface vs. concave; first caudal with flat ventral surface
vs. median groove; first caudal neural spine posteriorly sloped vs.
vertical; pubic shaft straight vs. strongly posteriorly curved;
transition between anterior pubic shaft and dorsal edge of pubic foot
rounded vs. angled; anterior pubic foot more pointed; obturator process
less prominent; ischial foot limited to anterior expansion vs. expanded
some posteriorly; ischiopubic ratio 82% vs. 92%. However, it seems the
authors depended on the literature for their information on Archaeornithomimus
(e.g. thinking the ischia are AMNH 21798 instead of 6558, and that the
first caudal would be amphicoelous and grooved because Smith and Galton
said in the proximal caudal paragraph "The centra are amphicoelous ...
and each has a shallow ventral groove"), and this led to misinformation
and incomplete statements. Regarding size, there is an ilium ~380 mm
long in AMNH 6570, larger than even LH-02-01. The proximal five
caudals of AMNH 21790 all lack median grooves ventrally and the
posterior articular surface of the first centrum is still attached to a
broken off part of the second centrum so cannot be evaluated. The
first caudal neural spine has been broken away since Smith and Galton's
drawing, and the shape of the other neural spines differ from the
drawing in greater to lesser degrees. There is a distal pubis in AMNH
6570 with an almost identical foot to LH-02-01, including the rounded
anterodorsal edge, more pointed anterior foot and shorter posterior
foot. Finally, there is no indication pubes AMNH 21799 and ischia AMNH
6558 were associated, and indeed five right proximal tibiae are known
from site 140 that can differ in size ~10% from each other. Thus sacra
AMNH 21790 and LH-02-01 may be distinct based on central lengths, pubes
AMNH 21799 and LH-02-01 / AMNH 6570 (in part) seem to be distinct, as
do ischia AMNH 6558 and LH-02-01. However, undescribed pubes AMNH
21798 are intermediate in having a slight anterior bow, rounded
anterodorsal transition, less pointed anterior foot and intermediate
posterior foot length. This suggests the sacral and ischial
differences could easily be due to the low sample size (N = 2) of each
element and that considering all of these specimens to be conspecific
is the most realistic and functional conclusion. Since the lectotype
is a partial pes not comparable to LH-02-01 at all (and without any
proposed diagnosis or comparison to other Iren Dabasu ornithomimosaur
pedal specimens), the other rational option would be to limit Archaeornithomimus asiaticus
to AMNH 6565 and potentially other comparable specimens (e.g. AMNH
6568, 21616, 30240B/C and parts of 6570 and 6576) while leaving the
non-pedal specimens as Ornithomimosauria indet.. Alternatively,
LH-02-01 and e.g. AMNH 21790 or 6558 could be given names, but again
the vast majority of specimens would be incomparable and relegated to
Ornithomimosauria indet.. Given this data and pending the description
of some of the multitude of unpublished specimens, all Iren Dabasu
ornithomimosaurs are listed here under Archaeornithomimus asiaticus.
(see page for references)
unnamed therizinosaurid (Gilmore, 1933)
Middle-Late Campanian, Late Cretaceous
AMNH 138,
Iren Dabasu Formation, Inner Mongolia, China
Material- (AMNH 6368; syntype of Alectrosaurus olseni) humerus
(390 mm), manual ungual I (190 mm on curve), manual phalanx II-1 (74 mm)
?...
(AMNH 21784) three mid caudal vertebrae (~41, ~38, ~35 mm), distal caudal centrum (~24 mm) (Mader and Bradley, 1989)
?... (uncollected) two or three elements (Mader and Bradley, 1989)
Comments- AMNH 6368 was found on May 4 1923 at Third Asiatic Expedition field site 138, 30 meters away from the lectotype of Alectrosaurus
olseni and was thought to belong to the same individual in the field.
Gilmore (1933) prefered to treat them as two individuals. He placed them in
the same species due to the similarity between the manual unguals, which are
“laterally compressed, strongly curved and have sharply curved extremities”,
as well as the association in the field and slenderness. Gilmore originally diagnosed
A. olseni partially on the characteristics of this specimen. Rozhdestvensky (1970) recognized the similarity to Therizinosaurus and referred Alectrosaurus to Therizinosauridae, but Barsbold (1976) and Perle (1977) correctly removed the forelimb from that taxon based on supposed Alectrosaurus
specimen IGM 100/50 with small humerus and manual ungual. Mader
and Bradley (1989) described the specimen in detail as a segnosaurid.
Zanno (2010) notes the slender proportions, poorly defined medial
tuberosity, and posterior humeral trochanter indicates it is not
referable to Neimongosaurus, and it is more gracile than Erliansaurus and lacks
that genus' distinctive crest-shaped posterior trochanter. Zanno (2006) recovered it as more derived than Alxasaurus in her phylogenetic analysis, and most recently Hartman et al. (2019) found it sister to Segnosaurus in Therizinosauridae.
AMNH 21784 was also discovered on May 4 1923 at Third Asiatic Expedition
field site 138, but
not catalogued until 1984 and not described until 1989 by Mader and
Bradley. While those authors described them as Theropoda incertae
sedis, they may be therizinosauroid based on their resemblence to Alxasaurus'
mid caudals (e.g. short centra that become shorter distally; low neural
spines; similarly placed transverse processes) although their neural
spines are more posteriorly restricted and their prezygapophyses are
longer. Although Mader and Bradley considered the caudals too small to
belong to the same individual as the humerus, they were comparing them
to theropod proximal caudals as they were unaware of therizinosauroids'
modified caudal series with relatively homogenous caudals decreasing in
length distally. In fact, given that Alxasaurus'
holotype humerus is 4% longer than AMNH 6368, its fourteenth-eighteenth
caudals are also slightly longer (39-44 mm) as is an isolated distal
caudal (26 mm), so the sizes are appropriate to belong to a single
therizinosauroid individual. As they are from the same field site,
that is provisionally accepted here. Mader and Bradley also noted "two
or three unspecified [theropod] elements that regrettably were not
collected due to their "poor condition" (Granger's field record, Third
Asiatic Expedition)" found with the forelimb and caudal material and
might also belong to the same individual.
References- Gilmore, 1933. On the Dinosaurian Fauna of the Iren Dabasu
Formation. Bulletin American Museum of Natural History. 67, 23-78.
Rozhdestvensky, 1970. Giant claws of enigmatic Mesozoic reptiles. Paleontological
Journal. 1970(1), 131-141.
Barsbold, 1976. New data on Therizinosaurus (Therizinosauridae, Theropoda). In Kramarenko, Luvsandansan, Voronin, Barsbold, Rozhdestvensky, Trofimov and Reshetov (Eds.).
Paleontology and Biostratigraphy of Mongolia. The Joint Soviet-Mongolian
Paleontological Expedition, Transactions. 3, 76-92.
Perle, 1977. On the first discovery of Alectrosaurus
(Tyrannosauridae, Theropoda) from the Late Cretaceous of Mongolia. Problemy
Geologii Mongolii. 3, 104-113.
Mader and Bradley, 1989. A redescription and revised diagnosis of the syntypes
of the Mongolian tyrannosaur Alectrosaurus olseni. Journal of Vertebrate
Paleontology. 9(1), 41-55.
Zanno, 2006. The pectoral girle and forelimb of the primitive therizinosauroid
Falcarius utahensis (Theropoda, Maniraptora): Analyzing evolutionary
trends within Therizinosauroidea. Journal of Vertebrate Paleontology. 26(3),
636-650.
Zanno, 2010. A taxonomic and phylogenetic re-evaluation of
Therizinosauria (Dinosauria: Maniraptora). Journal of Systematic
Palaeontology. 8(4), 503-543.
Hartman, Mortimer, Wahl, Lomax, Lippincott and Lovelace, 2019. A new
paravian dinosaur from the Late Jurassic of North America supports a
late acquisition of avian flight. PeerJ. 7:e7247.
undescribed Therizinosauroidea (Dong, 1992)
Middle-Late Campanian, Late Cretaceous
Iren Dabasu Formation, Inner Mongolia, China
Material- (Erenhot Dinosaur Museum coll.) dentary, teeth (Currie and Eberth, 1993)
(IVPP coll.) (isolated) many limb elements (Dong, 1992)
(IVPP and PIN coll.) tens of specimens (Cuirrie and Eberth, 1993)
Comments- Currie and Eberth
(1993) stated "A rough tally of Sino-Soviet field
identifications shows that ... 'theropods' (including large theropods,
small theropods and segnosaurs, but not ornithomimids) were more common
(400 specimens)" and that "The apparent high numbers of carnivorous
dinosaurs can be attributed mostly to ornithomimids [>1000 elements]
and segnosaurids" indicating some significant number of those 400
'theropods' found in June-July 1959 were therizinosaurs.
Dong (1992) reported "In July 1988, the expedition of the CCDP came to
Erenhot (Fig.85) where they collected ... Many limb bones [which] might
be identified as segnosaurs" and listed Segnosaurus
sp. as being present in the formation. He later (1993) said "at least
two taxa of segnosaurs" were represented. Currie and Eberth (1993)
stated "isolated elements were commonly recovered by the Sino-Canadian
expeditions in 1988 and 1990. A well-preserved dentary with teeth is in
the collections of the Erenhot Dinosaur Museum. The isolated elements
are indistinguishable from Erlicosaurus andrewsi and Segnosaurus ghalbiensis
bones in the collections of the Paleontological Institute (at the
Central State Museum) in Ulaan Baatar. There are a few elements from
the Iren Dabasu that may also be referable to the more poorly known
segnosaur Enigmosaurus." They listed both Segnosaurus sp. and Erlikosaurus
sp. as present, but given the poor state of knowledge of therizinosaur
diversity in the early 90s, they may actually belong to the
contemporaneous Erliansaurus,
Neimongosaurus, and/or the
taxon to which the forelimb AMNH 6368 belongs instead. Precise
localities have not been published, but the CCDP excavated several
(Currie and Eberth, 1993: Table 3), all around Iren Nor and far from
Sanhangobi where Erliansaurus and Neimongosaurus were found.
References- Dong, 1992. Dinosaurian Faunas of China: China Ocean Press. 188 pp.
Currie and Eberth, 1993. Palaeontology, sedimentology and palaeoecology of the
Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia, People’s Republic
of China. Cretaceous Research. 14, 127-144.
Dong, 1993. The field activities of the Sino-Canadian Dinosaur Project
in China, 1987-1990. Canadian Journal of Earth Sciences. 30(10),
1997-2001.
Kuszholia sp. (Yao, Wang, Sullivan, Wang, Stidham and Xu, 2015)
Middle-Late Campanian, Late Cretaceous
Iren Dabasu Formation, Inner Mongolia, China
Material- (IVPP V20377) anterior dentaries (symph 8.0 mm)
Comments-
This was discovered in 2012 at "a rare microvertebrate locality within
the Iren Dabasu Formation, about 16 km northeast of Erenhot City",
which would put it in localities Q-T of Xing et al. (2012). Yao et al.
(2015) referred IVPP V20377 to Caenagnathasia
sp., and it does possess all characters here listed as diagnostic for
that taxon. Of their characters listed as varying between Caenagnathasia specimens, most also vary between Chirostenotes
specimens (posterior surface of symphysis with tubercle; chin-like
eminence between anterior and ventral surfaces; pneumatic foramen in
front of mandibular fenestra on lateral surface of dentary; depression
on posteroventral margin of symphysis [ontogenetic?]), while paired
second anterior occlusal grooves flanking first anterior occlusal
groove is polymorphic in the holotype, and lateral projections on
lingual ridges may be absent in ZIN PH 2354/16 due to preservation.
This leaves presence of a median symphyseal groove on the posterodorsal
depression in the holotype and IVPP V20377 but not ZIN PH 2354/16
(ontogenetic?) and on the posteroventral symphysis of IVPP V20377 but
not the holotype or ZIN PH 2354/16 (taxonomic?). The age difference
suggests the specimen is not conspecific with the Bissekty species.
References-
Xing, He, Li and Xi, 2012. A review on the study of the stratigraphy,
sedimentology, and paleontology of the Iren Dabasu Formation, Inner
Mongolia. In Dong (ed.). Proceedings of the Thirteenth Annual Meeting
of the Chinese Society of Vertebrate Paleontology. China Ocean Press.
1-44.
Yao, Wang, Sullivan, Wang, Stidham and Xu, 2015. Caenagnathasia
sp. (Theropoda: Oviraptorosauria) from the Iren Dabasu Formation (Upper Cretaceous:
Campanian) of Erenhot, Nei Mongol, China. Vertebrata PalAsiatica. 53(4), 291-298.
Wang, Zhang and Yang, 2018. Reevaluation of the dentary structures of
caenagnathid oviraptorosaurs (Dinosauria, Theropoda). Scientific
Reports. 8:391.
Avimimus sp. nov. (Makovicky, 1995)
Middle-Late Campanian, Late Cretaceous
Iren Dabasu Formation, Inner Mongolia, China
Material- (AMNH 6570 in part; paratype of Ornithomimus asiaticus) fibula (Chiappe, Norell and Clark, 2002)
(AMNH 6576 in part; paratype of Ornithomimus asiaticus) proximal caudal vertebra (Makovicky, 1995)
?(AMNH 6754) distal metatarsal III (Ryan, Currie and Russell, 2001)
?..?(AMNH
6755; 'AMNH 6555' of Funston, Currie, Ryan and Dong, 2019) incomplete
metatarsus (mtII 100, mtIV 99 mm) (Ryan, Currie and Russell, 2001)
(AMNH 25569) caudal vertebra, ten vertebrae, three phalanges, five unguals including pedal ungual (~18 mm) (AMNH online)
(AMNH coll.) dorsal vertebrae (Makovicky, 1995)
?(IVPP V16313.a) manual ungual ?I (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16314; = TMP 1992.302.0102) proximal tarsometatarsus (Ryan, Currie and Russell, 2001)
?...(IVPP V16341) tarsometatarsus (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16315) metatarsal II (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16316.a) pedal ungual III (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16316.b) pedal ungual III (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16316.c; = TMP 1992.302.0119A) pedal ungual II/IV (Ryan, Currie and Russell, 2001)
(IVPP V16316.d; = TMP 1992.302.0119B) pedal ungual II/IV (Ryan, Currie and Russell, 2001)
(IVPP V16317.a) incomplete proximal caudal vertebra (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16317.b) mid caudal vertebra (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16318; ?= TMP 1992.302.0344) (juvenile) posterior cervical vertebra (Ryan, Currie and Russell, 2001)
(IVPP V16318.a) incomplete anterior dorsal vertebra (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16318.b; ?= IVPP 160788-124) incomplete posterior dorsal vertebra (Makovicky, 1995; Funston, Currie, Ryan and Dong, 2019)
(IVPP V16319) some of "IVPP V163.." below (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16320) (<1 year old juvenile) distal tibiotarsus (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16321) proximal metatarsal II (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16322.a; = TMP 1992.302.0150) proximal tibia (Ryan, Currie and Russell, 2001)
(IVPP V16322.b) proximal tibia (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16322.c) proximal tibia (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16323.a) third dorsal vertebra (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16324) some of "IVPP V163.." below (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16325) some of "IVPP V163.." below (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16326) metatarsal II (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16327; = TMP 1992.302.0116) partial scapulocoracoid (Ryan, Currie and Russell, 2001)
(IVPP V16328) (juvenile) mid sacral centrum (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16329.a; = IVPP 180788-123) last or penultimate cervical vertebra (Makovicky, 1995)
(IVPP V16329.b) second dorsal vertebra (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16330; = IVPP 160788-122) posterior sacral fragment (Makovicky, 1995)
(IVPP V16331) some of "IVPP V163.." below (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16332.a; = TMP 1992.302.0140) partial second dorsal vertebra (Ryan, Currie and Russell, 2001)
(IVPP V16332.b) partial first dorsal vertebra (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16333) some of "IVPP V163.." below (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16334.a; = TMP 1992.302.0149) proximal femur (Ryan, Currie and Russell, 2001)
(IVPP V16335.a) (juvenile) distal tarsal IV fused to proximal metatarsal IV (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16335.b) distal metatarsal II/III
....(IVPP V16335.c) distal metatarsal II/III
....(IVPP V16336) distal metatarsal IV
(IVPP V16337) (>2 year old adult) distal tarsometatarsus (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16338; = TMP 1992.302.0110) distal femur (Ryan, Currie and Russell, 2001)
(IVPP V16339) some of "IVPP V163.." below (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16340; = TMP 1992.302.0117) proximal humerus (Ryan, Currie and Russell, 2001)
?(IVPP V16342; = TMP 1992.302.0104) partial frontal (Ryan, Currie and Russell, 2001)
(IVPP V16343) distal metacarpal I (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16344) some of "IVPP V163.." below (Funston, Currie, Ryan and Dong, 2019)
(IVPP V16345) some of "IVPP V163.." below (Funston, Currie, Ryan and Dong, 2019)
(IVPP V163...) four dorsal vertebrae, distal metatarsal, pedal ungual
III, eight pedal unguals II/IV (Makovicky, 1995; Funston, Currie, Ryan
and Dong, 2019)
(PIN coll.) material (Currie and Eberth, 1993)
Diagnosis- (after Funston et al., 2019) three cervicodorsal vertebrae (unknown in A. nemegtensis); deeper femoral intercondylar groove.
Comments-
Makovicky (1995) stated "Avimimid vertebrae were collected by the
American Museum Central Asian expedition in 1922 at Iren Dabasu.
However, they were not recognised as such, and were catalogued with Ornithomimus asiaticus (now Archaeornithomimus asiaticus) material", but the Archaeornithomimus
type material was discovered in 1923 including a proximal caudal
referenced by the author. While he says "one of the caudals collected
by the American Museum's Central Asiatic Expeditions (AMNH 6576), [has]
a small pointed tubercle separates the ventromedial corners of the
chevron facets" this specimen number includes almost a hundred elements
from the Johnson Quarry AMNH locality 141 generally referred to Archaeornithomimus.
A fibula (AMNH 6570) was figured as "alvarezsaurid fibula from Iren
Dabasu (Inner Mongolia, China)" by Chiappe et al. (2002), but Longrich
and Currie (2009) stated "it more closely resembles the fibula of
Avimimidae, which are common at this locality (N.R.L., pers. obs.)."
Again this number includes over two hundred paratype Archaeornithomimus
elements, this time from the Kaisen Quarry AMNH locality 140.
Makovicky also stated "A large number of dorsal vertebrae are present
in the American Museum ... collections from Iren Dabasu", which are
likely to also be catalogued under AMNH 6570 and/or 6576. Longrich's
claim is supported here however as the fibula has an anteriorly
projected iliofibularis tuber as in Avimimus
but unlike the laterally directed tuber of e.g. IGM 100/99 and is less
reduced distally in anteroposterior width than the latter. A distal
caudal
(AMNH coll.) described by Makovicky "has the morphology of a typical
coelurosaurian
distal caudal, but is otherwise undiagnostic. The possibility that it
may originate from an avimimid is suggested by its small size. It
should be noted, however, that it could just as conceivably be from the
tail of a juvenile Archaeornithomimus from the same bonebed." Indeed, the A. nemegtensis bonebed show Avimimus has short distal caudals like other caenagnathoids and unlike ornithomimosaurs, so this specimen is here referred to Archaeornithomimus.
The AMNH online catalogue lists AMNH 25569 as "10 vertebrae 1 caudal
vertebra 3 phalanges & 5 claws" of Saurischia from the Johnson
Quarry with a pedal ungual photographed as Avimimidae. A metatarsus
(AMNH 6755) and third metatarsal (AMNH 6754) were listed on the
museum's online catalogue as Elmisaurus sp., but also referred to Avimimus by Ryan et al. (2001) and Funston et al. (for AMNH 6755 at least).
Ryan et al.'s poster indicated both were found in 1923 and were
possibly associated. AMNH 6755 does seem smaller than other fused Avimimus
and less slender, so further study is necessary. Note Funston et al.
(2019) describe and figure this as AMNH 6555, which is the number of
ornithischian material. Funston et al. state the main Iren Dabasu Avimimus
bonebed "was originally discovered by a Sino-Soviet expedition in 1959,
which used bulldozers to excavate the site. It was revisited in 1987
and 1988 by the Sino-Canadian expedition and numerous fragmentary bones
representing all regions of the skeleton (Fig. 2) were recovered from
the spoil piles left by the Sino-Soviet bulldozers. Unfortunately, the
material collected by the Sino-Soviet expedition still awaits
preparation and it may never be available for study." This indicates
it must be locality K of Currie and Eberth (1993), which was CCDP
locality 1 and may correspond to AMNH locality 141. One element from
this excavation was described and figured by Kurzanov (1987) as "a left
avimimid femur from the Upper Cretaceous Iren-Nor locality in China
(specimen PIN, no. 2549-100)" (translated) but is here placed in
Oviraptoridae. While the PIN Sino-Soviet Avimimus
material remains undescribed (although mentioned by Currie and Eberth),
the Sino-Canadian material was noted by Dong et al. (1989) who reported
"seven pedal elements of
Avimimus" discovered in July 1988 and Dong (1992) refers to
"fused tarsometatarsi of Avimimus" recovered on that expedition in July 1998.
Currie and Eberth (1993) stated "Direct comparison between isolated Avimimus bones from the Iren Dabasu and the type specimen of Avimimus portentosus
in Moscow failed to reveal any differences" and said the "material is
presently under review (Currie, Zhao and Kurzanov, in preparation)."
The Sino-Canadian Avimimus material was eventually described by Makovicky, Ryan et al., then officially by Funston et al. as Avimimus
sp.. Makovicky used IVPP field numbers (of which IVPP 180788-123 may
be a mistake for IVPP 160788-123 to better match the other two listed
numbers and only differ in the last digit), Ryan et al. used TMP
numbers in their poster, and the specimens were seemingly eventually
transported back to the IVPP for permanent storage. Note the "isolated
left frontal" mentioned by Sues et al. is not described in Funston et
al. (although the frontal is highlighted as preserved in their figure
2), but Funston (2019) describes it in his thesis chapter that was
developed into that paper. In it he says "whether it pertains to an
avimimid or another oviraptorosaur is uncertain. It can be
distinguished from other theropods by the large, incising nasal
contact, which is similar to the morphology of Elmisaurus rarus",
so it's possible this belonged to the Iren Dabasu caenagnathid taxon
represented by dentaries IVPP V20377. IVPP 160788-124 is one of three
posterior dorsals without a ventral keel, so may be IVPP V16318.b which
was the only one of these figured by Funston et al.. Another tricky
specimen is TMP 1992.302.0344 as illustrated in Ryan et al.'s poster,
which is tentatively identified as posterior cervical IVPP V16318 as it
seems to have a ventrally placed parapophysis, an elongate centrum with
vertical articular surfaces, two or three central foramina and a dark
neurocentral boundary perhaps representing an open suture, but contra
to Funston et al.'s description is not notably smaller than other
cervicals. While Funston et al. only referred to the Iren Dabasu
material as Avimimidae gen. et sp. indet., they also stated "the
cervicodorsal vertebrae differ in number (three with hypapophyses) from
those of Avimimus portentosus (MPC-D 100/129), although cervicodorsal number in Avimimus nemegtensis
is unknown. The distal condyles of the femur (Fig. 6) are separated
much more deeply than is typical in avimimids, and metatarsals II and
IV (Fig. 8) are much more disparate in size", but ended up concluding
"the available material from the Iren Dabasu bonebed is too incomplete
to confidently erect a new taxon, but future
preparation of the Russian material (or collection of new material) may
result in its taxonomic distinction from other avimimids."
References- Kurzanov, 1987. Avimimidae and the problem of the origin
of birds. Trudy, Sovmestnaa Sovetsko-Mongolskaa paleontologiceskaa
ekspedicia. 31, 1-95.
Dong, Currie and Russell, 1989. The 1988 field program of the Dinosaur Project. Vertebrata Palasiatica. 27(3), 233-236.
Dong, 1992. Dinosaurian Faunas of China. China Ocean Press. 188 pp.
Currie and Eberth, 1993. Palaeontology, sedimentology and palaeoecology of the
Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia, People's Republic
of China. Cretaceous Research. 14, 127-144.
Makovicky, 1995. Phylogenetic aspects of the vertebral morphology of Coelurosauria
(Dinosauria: Theropoda). Masters thesis, University of Copenhagen. 311 pp.
Ryan, Currie and Russell, 2001. New material of Avimimus portentosus
(Theropoda) from the Iren Dabasu Formation (Upper Cretaceous) of the
Erenhot region of Inner Mongolia. Journal of Vertebrate Paleontology.
21(3), 95A.
Chiappe, Norell and Clark, 2002. The Cretaceous, short-armed Alvarezsauridae,
Mononykus and its kin. In Chiappe and Witmer (eds.). Mesozoic
Birds: Above the Heads of Dinosaurs. University of California Press. 87-120.
Longrich and Currie, 2009 (online 2008). Albertonykus borealis, a new alvarezsaur (Dinosauria:
Theropoda) from the Early Maastrichtian of Alberta, Canada: Implications for
the systematics and ecology of the Alvarezsauridae. Cretaceous Research. 30(1),
239-252.
Funston, 2019. Anatomy, systematics, and evolution of Oviraptorosauria
(Dinosauria, Theropoda). PhD thesis, University of Alberta. 774 pp.
Funston, Currie, Ryan and Dong, 2019. Birdlike growth and mixed-age
flocks in avimimids (Theropoda, Oviraptorosauria). Scientific Reports.
9:18816.
 |
| Small portion of material catalogued under AMNH 6570 and assigned to Archaeornithomimus. Note the ungual in the upper left is not ornithomimid and here compared favorably to Citipati. My photo courtesy of the AMNH. |
cf. Citipati (Kurzanov, 1987)
Middle-Late Campanian, Late Cretaceous
Iren Dabasu Formation, Inner Mongolia, China
Material- ?(AMNH 6570 in part; paratype of Ornithomimus asiaticus) partial manual ungual ?I (~39 mm) (pers. obs.)
(PIN 2549-100) femur (~160 mm) (Kurzanov, 1987)
Comments- Kurzanov (1987) briefly described and figured "a
left avimimid femur from the Upper Cretaceous Iren-Nor locality in
China (specimen PIN, no. 2549-100)" (translated), which Currie and
Eberth (1993) indicate was found by the Sino-Soviet expedition, which means it was collected between June 14 and July
17 at their localities K (= AMNH locality 141?), L or P. Kurzanov
referred it to Avimimidae based on the "accessory condyle" (which is
just the lateral condyle being separated from the ectocondylar tuber by
a fibular groove as in most theropods), the broad intercondylar flexor
groove and similarities of the trochanteric crest. Indeed, he stated "the
only not very significant difference is expressed in the fusion of the
large and small trochanters, while in Avimimus they are separated by a
narrow gap." On the other hand, Osmolska (1996) stated "There is a
great resemblance between the femur in [Bagaraatan] ostromi and the femur PIN
2549-100" in that "Both femora have similarly shaped proximal and
distal ends, ... well pronounced articular heads and femoral necks, the
poorly delimited lesser trochanters, which are as high as the greater,
and in the presence of the protuberances on the lateral surface."
Additionally, "The distal ends of femora are also similarly shaped in
both compared forms" with "distinctive tibiofibular crests ('condylus
lateralis' in Kurzanov 1987)." While PIN 2549-100 is similar in shape
to both Iren Dabasu Avimimus and Bagaraatan, the latter both have
accessory trochanters (usually misidentified as a large, distally
placed anterior trochanter in Avimimus) which are absent in PIN
2549-100, Avimimus differs from PIN 2549-100 and Bagaraatan in lacking
a distal ectocondylar notch defining the tuber, Bagaraatan differs from
PIN 2549-100 and Avimimus in having a narrow flexor groove, and PIN
2549-100 differs from at least Avimimus in having a fourth trochanter
reduced to "a
slight roughness, located almost under the head of the femur on its
medial side" (Kurzanov, 1987) (unknown in Bagaraatan). Currie and
Eberth (1993) believed PIN 2549-100 "is probably from a troodontid" and
"provisionally referred to Saurornithoides", but Averianov and Sues
(2012) concluded it "is probably troodontid but cannot be definitely
referred to Saurornithoides" and "should be listed as Troodontidae
indet." However, scoring this in Hartman et al.'s maniraptoromorph analysis results in identical scorings to Citipati osmolskae, with one more step needced to move it sister to Avimimus and two more to move it to Troodontidae (as sister to Linhevenator). Given the stratigraphic and geographic proximity, it is provisionally assigned to cf. Citipati here pending description of the holotype's femur (IGM 100/979 and 1004 have crushed and poorly exposed femora).
The hypodigm of Archaeornithomimus asiaticus includes two collections
of largely undescribed and unassociated specimens, AMNH 6570 from Third
Asiatic Field site 140 and AMNH 6576 from site 141, discovered between April 22 and May 25 1923. Based on personal
examination (July 2009), multiple elements in these collections do not
belong to Archaeornithomimus, among which is an ungual in a box of
phalanges and calcanea under AMNH 6570. It is moderately curved with a
large, proximally placed flexor tubercle and resembles both
Sinornithoides' pedal ungual I and Citipati's
manual ungual I in its
preserved portion. As it is twice the length of Iren Dabasu's
troodontid pedal unguals I but scales well to PIN 2549-100, the latter
identification is provisionally preferred here.
References- Kurzanov, 1987. Avimimidae and the problem of the origin
of birds [in Russian]. Trudy, Sovmestnaa Sovetsko-Mongolskaa paleontologiceskaa
ekspedicia. 31, 1-95.
Currie and Eberth, 1993. Palaeontology, sedimentology and
palaeoecology of the Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia,
People's Republic of China. Cretaceous Research. 14, 127-144.
Osmolska, 1996. An unusual theropod dinosaur from the Late Cretaceous Nemegt
Formation of Mongolia. Acta Palaeontologica Polonica. 41, 1-38.
Averianov and Sues, 2012. Correlation of Late Cretaceous continental vertebrate
assemblages in middle and central Asia. Journal of Stratigraphy. 36(2), 462-485.
undescribed Troodontidae (Currie and Eberth,
1993)
Middle-Late Campanian, Late Cretaceous
Erenhot, Iren Dabasu Formation, Inner Mongolia, China
Material- (AMNH 6570 in part; paratype of Ornithomimus asiaticus) (juvenile or subadult) axis, third cervical vertebra, fifth cervical vertebra (Makovicky,
1995)
?(AMNH 6576 in part; paratype of Ornithomimus asiaticus) pedal ungual I (~17 mm) (pers. obs.)
(AMNH 21751) distal metatarsals III (Currie and Eberth,
1993)
(AMNH 21772) metatarsal II (~183 mm) (Currie and Eberth, 1993)
(AMNH 25570) three vertebrae (AMNH online)
(AMNH 30261) proximal metatarsal (AMNH online)
(AMNH 30262) proximal tibial fragment (AMNH online)
(AMNH 30263) proximal tibial fragment (AMNH online)
(AMNH 30264) tibial fragment, fibular fragment (AMNH online)
(AMNH 30265) proximal tibia (AMNH online) 142
(AMNH 30266) proximal fibula (AMNH online) 142
(AMNH 30267) proximal tibia (AMNH online)
(AMNH 30268) proximal fibula (AMNH online)
(AMNH 30269) proximal fibula (AMNH online) 142
(AMNH 30270) proximal fibula (AMNH online)
(AMNH 30271) partial astragalus (AMNH online)
(AMNH 30272) partial astragalus (AMNH online)
(AMNH 30273) partial astragalus (AMNH online)
(AMNH 30274) partial astragalus (AMNH online)
(AMNH 30275) distal humerus (AMNH online)
(AMNH 30276) distal humerus (AMNH online)
(AMNH 30277) distal humerus (AMNH online)
(AMNH 30278) proximal humerus (AMNH online)
(AMNH 30279) proximal humerus (AMNH online)
(AMNH 30280) proximal ulna (AMNH online)
(AMNH 30281) distal radius (AMNH online)
(AMNH 30282) distal radius (AMNH online)
(AMNH 30283) proximal scapula (AMNH online)
(AMNH 30284) proximal scapula (AMNH online)
(AMNH 30285) scapular blade (AMNH online)
(AMNH 30286) pedal ungual I (~19 mm) (AMNH online)
(AMNH 30287) proximal manual ungual (AMNH online)
(AMNH 30288) two posterior cervical or proximal caudal vertebrae (AMNH online)
(AMNH 30289) distal metatarsal IV (AMNH online)
(AMNH 30290) distal metatarsal IV (AMNH online)
(AMNH 30291) distal metatarsal IV (AMNH online)
(AMNH 30292) distal metatarsal III (AMNH online)
(AMNH 30293) distal metatarsal III (AMNH online)
(AMNH 30294) distal metatarsal II (AMNH online)
(AMNH 30295) distal metatarsal II (AMNH online)
(AMNH 30296) distal metatarsal II (AMNH online)
(AMNH 30297) distal metatarsal II (AMNH online)
(AMNH 30300) partial ilium (AMNH online) 142
(AMNH 30301) proximal ?pubis (AMNH online) 142
(AMNH 30302) ?ilial fragment (AMNH online)
(AMNH 30303) partial synsacrum (AMNH online) 142
(AMNH 30304) proximal ?pubis (AMNH online)
?(AMNH 30305) last sacral vertebra (~25 mm) (AMNH online)
(AMNH 30306) partial synsacrum (AMNH online)
(AMNH 30307) synsacral fragment(AMNH online)
(AMNH 30308) partial posterior cervical vertebra (AMNH online)
(AMNH 30309) partial posterior cervical vertebra (AMNH online)
(AMNH 30310) partial anterior dorsal vertebra (AMNH online)
(AMNH 30311) partial anterior dorsal vertebra (AMNH online)
(AMNH 30312) partial anterior dorsal centrum (AMNH online)
(AMNH 30313) incomplete anterior dorsal centrum (AMNH online)
(AMNH 30314) partial anterior dorsal centrum (AMNH online)
(AMNH 30315) incomplete anterior dorsal centrum (AMNH online)
(AMNH 30316) partial anterior dorsal vertebra (AMNH online)
(AMNH 30317) incomplete anterior dorsal centrum (~24 mm) (AMNH online)
(AMNH 30318) anterior dorsal centrum (~26 mm) (AMNH online)
(AMNH 30320) anterior dorsal centrum (~26 mm) (AMNH online) 142
(AMNH 30321) partial anterior dorsal vertebra (~26 mm) (AMNH online)
?(AMNH 30322) anterior dorsal centrum (~32 mm) (AMNH online)
(AMNH 30323) incomplete posterior dorsal centrum (~22 mm) (AMNH online)
(AMNH 30324) incomplete dorsal centrum (AMNH online)
(AMNH 30325) posterior dorsal centrum (~26 mm) (AMNH online)
(AMNH 30326) posterior dorsal centrum (~27 mm) (AMNH online)
(AMNH 30327) posterior dorsal centrum (~27 mm) (AMNH online)
(AMNH 30328) incomplete posterior dorsal vertebra (~27 mm) (AMNH online)
(AMNH 30329) incomplete posterior dorsal vertebra (~29 mm) (AMNH online)
(AMNH 30330) proximal caudal centrum (~23 mm) (AMNH online)
(AMNH 30336) ?central fragment (AMNH online)
(AMNH 30337) incomplete distal caudal vertebra (~35 mm) (AMNH online)
(AMNH 30338) mid caudal vertebra (~32 mm) (AMNH online)
(AMNH 30339) incomplete mid caudal vertebra (~31 mm) (AMNH online)
(AMNH 30340) incomplete mid caudal vertebra (AMNH online)
(AMNH 30341) partial distal caudal vertebra (AMNH online)
(AMNH 30342) mid caudal vertebra (~29 mm) (AMNH online)
(AMNH 30343) mid caudal vertebra (~30 mm) (AMNH online)
(AMNH 30344) distal caudal vertebra (~32 mm) (AMNH online)
(AMNH 30345) incomplete mid caudal vertebra (~25 mm) (AMNH online)
(AMNH 30346) partial distal caudal vertebra (AMNH online)
(AMNH 30347) partial distal caudal vertebra (AMNH online)
(AMNH 30348) distal caudal vertebra (~31 mm) (AMNH online)
(AMNH 30349) fragmentary distal caudal vertebra (AMNH online)
(AMNH 30350) distal caudal vertebra (~33 mm) (AMNH online)
(AMNH 30351) incomplete distal caudal vertebra (~31 mm) (AMNH online)
(AMNH 30352) partial distal caudal vertebra (AMNH online)
(AMNH 30353) incomplete distal caudal vertebra (AMNH online)
(AMNH 30354) distal caudal vertebra (~23 mm) (AMNH online)
?(AMNH 30355) mid caudal vertebra (~32 mm) (AMNH online)
(AMNH 30356) proximal caudal centrum (~28 mm) (AMNH online)
(AMNH 30357) incomplete proximal caudal vertebra (~25 mm) (AMNH online)
(AMNH 30358) proximal caudal vertebra (~24 mm) (AMNH online)
(AMNH 30359) incomplete proximal caudal centrum (AMNH online)
(IVPP 230790-16; = IVPP 230090-16 of Currie and Eberth, 1993) metatarsal III (Currie and Eberth, 1993)
Comments-
Currie and Eberth (1993) stated "Troodontid bones are rare, but include
distinctive third metatarsals (AMNH 21751, 21772, IVPP 230090-16), in
which the distal articulation extends onto the posterior surfrace of
the bone in a broad tongue." However, Currie and Dong (2001) corrected
the identification of the second specimen, stating "AMNH 21772 is the
proximal end of a second metatarsal. It is identified as a troodontid
on the basis of its contact surface for the fourth metatarsal, its
size, and especially its lateromedial compression." The AMNH online
catalogue photo indicates most of the element is preserved and the
locality info is "8 mi. E. of station" indicating it was found in
localities 140-149 in 1923 or 1928. Currie and Dong describe AMNH
21751 as "two distal ends of third metatarsals [that] are about the
same size and represent left and right elements. Although they may
represent the same individual, the two fossils are different colours,
which suggests they may not have been found together." They indicate
these were "Collected in the 1920s by the third Central Asiatic
Expedition from exposures of the Iren Dabasu Formation (?Santonian)
near Erenhot." Currie and Dong list IVPP 230790-16 (presumably the
correct field number for Currie and Eberth's 'IVPP 230090-16') as a
metatarsal "Collected in 1990 from exposures of the Iren Dabasu
Formation (?Santonian) near Erenhot", which would make it found during
the second Sino-Canadian expedition. They state "the tongue-like
extensions of the third metatarsals from Iren Dabasu are flat like
those of Troodon ... , Borogovia ... , and Tochisaurus" but unlike the grooved surface of Sinornithoides or the distally restricted surface of Philovenator. This has since been identified in Bissekty Urbacodon sp. ZIN PH 2342/16, and it should be noted the extension of Tochisaurus is much shorter, while Mei, IGM 100/44, 100/140 and 100/1126 have a condition like Sinornithoides. Thus as hypothesized by Dong and Currie, at least AMNH 21751 and IVPP 230790-16 are closer to Troodon than Sinornithoides. Currie and Eberth stated "These bones are provisionally referred to Saurornithoides" (at the time a concept including Zanabazar)
without rationale, but Currie and Dong instead classified them as "an
unknown species of troodontid", stating they "cannot be identified
further without additional material."
Makovicky (1995) stated "A probable troodontid axis (AMNH 6570),
articulated with a third cervical vertebra, is present in the
collections of the American Museum of Natural History. This
identification is based on the morphology of the associated third
cervical and a probable fifth cervical, possibly from the same
individual, which strongly resembles those of Troodon.
The axis is from an immature individual as seen from absence of both
the odontoid and axial intercentrum." This specimen number includes
over two hundred paratype Archaeornithomimus
elements from the Kaisen Quarry AMNH locality 140, and the cervicals
described were not recognized in the material catalogued under it in
July 2009 (pers. obs.). However, a small ungual was noticed in AMNH
6576 (which includes almost a hundred paratype Archaeornithomimus
elements from the Johnson Quarry AMNH locality 141) that most closely
resembles a troodontid pedal ungual I in the slight curvature,
proximally placed flexor tubercle and posterodorsal extent being less
than its posteroventral extent.
The AMNH online catalogue lists AMNH 25570 as "Troodon ?",
consisting of "3 vertebrae." A large number of elements (AMNH
30261-30297, 30300-30318, 30320-30330, 30336-30359) are labeled are
labeled "Troodontid" on the AMNH online catalogue, each from the same
location ("8 mi. E. of station") and from AMNH Quarry 142 specifically
when visible in the photo (AMNH 30265, 30266, 32069, 30300, 30301,
30303, 30320). Given the similar preservation and number of elements
preserved, it is possible these represent two individuals, and that
several other specimens only identified to the level of Saurischia in
the online catalogue (AMNH 30245, 30247-30260, 30298-30299, 30360) that
are also from "8 mi. E. of station" may belong to them as well. Note
AMNH 30267 is incorrectly identified as a proximal fibula, while 30288
is called "Proximal end of metatarsal IV" but seems to be two vertebrae
instead, AMNH 30301 is called an "Ilium fragment." but may be a
proximal pubis (posterior edge downward in photo), AMNH 30302 is
labeled as "Acetabulum fragment." and indeed may be the ischial
peduncle and postacetabular base of a left ilium, AMNH 30304 is labeled
"Prox. end of ischium" but more closely resembles a proximal troodontid
pubis in the diverging peduncles and shallowly concave acetabular edge,
AMNH 30305 is a last sacral vertebra with a convex posterior central
face and 30322 is an anterior dorsal with convex anterior central face
so both may be alvarezsaurid instead. Scoring the material as
photographed in the online catalogue (with AMNH 30305 and 30322
excluded, and 30301 and 30304 interpreted as pubes) into Hartman et
al.'s maniraptoromorph matrix does result in it being troodontid, but
note examination of the specimens themselves would provide far more
data for each element and that it's currently only an assumption that
they belong to the same taxon.
References- Currie and Eberth, 1993. Palaeontology, sedimentology and
palaeoecology of the Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia,
People's Republic of China. Cretaceous Research. 14, 127-144.
Makovicky, 1995. Phylogenetic aspects of the vertebral morphology of Coelurosauria
(Dinosauria: Theropoda). Masters thesis, University of Copenhagen. 311 pp.
Currie and Dong, 2001. New information on Cretaceous troodontids (Dinosauria,
Theropoda) from the People's Republic of China. Canadian Journal of Earth Sciences.
38(12), 1753-1766.
Averianov and Sues, 2012. Correlation of Late Cretaceous continental vertebrate
assemblages in middle and central Asia. Journal of Stratigraphy. 36(2), 462-485.
 |
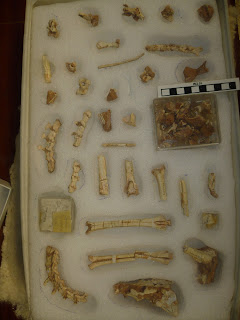
| Sample of unguals catalogued under AMNH 6576 as Archaeornithomimus. The bottom left one is not ornithomimid and is compared to pedal ugual I of Sinornithoides here, so suggested to be troodontid. Note too the juvenile ?Bactrosaurus pedal ungual in the box on the right. Scale = 100 mm. My photo courtesy of the AMNH. |
undescribed Dromaeosauridae (Gilmore, 1933)
Middle-Late Campanian, Late Cretaceous
Iren Dabasu Formation, Inner Mongolia, China
Material- (AMNH 6572) pedal phalanx II-1 (Ostrom, 1969)
(AMNH 21781) pedal ungual II (AMNH online)
?(IVPP 270790-4) tooth (~21x~9x? mm) (Currie and Zhao, 1993)
?(IVPP V16334.b) proximal femur (Funston, Currie, Ryan and Dong, 2019)
(IVPP coll.) teeth, elements (Dong, Currie and Russell, 1989)
(IVPP coll.) teeth and/or elements (Yao, Wang, Sullivan, Wang, Stidham and Xu, 2015)
Comments-
Gilmore (1933) noted the presence of "a few foot bones and other
fragmentary skeletal parts" of small theropods from the Iren Dabasu
Formation that he assigned to "Dromaeosaurinae Genus and species
indet.", but admitted that this assignment as opposed to Coeluridae or
Compsognathidae is based purely on their Cretaceous age. Thus while it
turned out at least two of these elements are apparently dromaeosaurid
(see below), Gilmore's rationale is equivalent to Maniraptora indet.
today and the fossils referenced plausibly included material now
recognized as avimimid, troodontid and ?alvarezsaurid. Ostrom (1969)
noted in a discussion of deinonychosaur pedal examples that "E. H.
Colbert has also discovered an isolated phalanx (AMNH 6572) in the
American Museum collections from the Iren Dabasu Formation of Mongolia
which compares almost exactly with the proximal phalanx of digit II of Deinonychus, but is perhaps 20 percent larger", which would make it somewhere around 46-52 mm long. He shows it questionably derived from Velociraptor
in his phylogram without explanation, which would not make sense in the
most recent interpretation of Iren Dabasu's age being contemporaneous
or slightly older than the Djadochta. Paul (1988) states "at the AMNH
is a hyper-extendable toe bone from the Late Cretaceous of Mongolia
that looks like a Velociraptor somewhat bigger than V. antirrhopus [= Deinonychus]",
but it is uncertain whether he saw it independant of Ostrom's text
(Paul, pers. comm. 6-2022). The AMNH online catalogue lists AMNH 21781
as an Iren Dabasu member of Dromaeosauridae represented by "Ungual of
pes (digit II)" and being found by Kaisen. Both AMNH 6572 and 21781
would have been found in Erenhot during the April 22 to May 25 1923
Central Asiatic Expedition.
Dong et al. (1989) state Velociraptor material was discovered in the July 1988 Sino-Canadian expedition to Erenhot, and Dong (1992) specifies "teeth of Velociraptor". Currie and Eberth (1993) state "Isolated dromaeosaurid teeth and bones are common in
the Iren Dabasu" and that "Most of these can be attributed to Velociraptor,
although some of the teeth suggest that there was a second, larger
species of an indeterminate dromaeosaurine dromaeosaurid." Yet no
rationale was presented, and the only two specified Iren Dabasu
dromaeosaurid elements in the literature are clearly not Velociraptor (AMNH 6572 is twice
the size, while IVPP 270790-4 is different from most dromaeosaurid
teeth as noted above). AMNH 6572 may belong to the supposed
dromaeosaurine though, based on size. Currie and Zhao (1993) figure
"Dromaeosaurid tooth (IVPP 270790-4) from
the Iren Dabasu Formation near Erenhot, People's Republic of China,
showing replacement pit on medial side of root", but given the slight
constriction basal to the
crown, convex distal edge and seeming lack of serrations, this may be
misidentified. Based on its similar field number to troodontid
metatarsal
IVPP 230790-16 it was probably also found in the Sino-Canadian
expedition of 1990.
Funston et al. (2019) notes a femoral head that
supposedly differs from Avimimus
in having fused anterior and greater trochanters, "which suggests that
it may be oviraptorid or, more likely, dromaeosaur." This is from the Avimimus
bonebed (locality K of Currie and Eberth [1993], which was CCDP
locality 1 and may correspond to AMNH locality 141) and recovered in
July 1988. Interestingly, femur PIN 2549-100 is from the same locality
and shares a trochanteric crest, but is here identified as
oviraptorid. It's possible they are from the same taxon (or even
individual).
Yao et al. (2015) note "small unarticulated bones and teeth, including
fossils of ... dromaeosaurids" from "a rare microvertebrate locality
within the Iren Dabasu Formation, about 16 km northeast of Erenhot
City."
References- Gilmore, 1933. On the dinosaurian fauna of the Iren Dabasu
Formation. Bulletin American Museum of Natural History. 67, 23-78.
Ostrom, 1969. Osteology of Deinonychus antirrhopus, an unusual theropod
from the Lower Cretaceous of Montana. Peabody Museum of Natural History Bulletin.
30, 1-165.
Paul, 1988. Predatory Dinosaurs of the World. Simon & Schuster. 464 pp.
Dong, Currie and Russell, 1989. The 1988 field program of The Dinosaur Project. Vertebrata PalAsiatica. 27(3), 233-236.
Dong, 1992. Dinosaurian Faunas of China. China Ocean Press. 188 pp.
Currie and Eberth, 1993. Palaeontology, sedimentology and palaeoecology of the
Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia, People s Republic
of China. Cretaceous Research. 14, 127-144.
Currie and Zhao, 1993 (published 1994). A new troodontid (Dinosauria, Theropoda) braincase from
the Dinosaur Park Formation (Campanian) of Alberta. Canadian Journal of Earth
Sciences. 30(10-11), 2234-2247.
Yao, Wang, Sullivan, Wang, Stidham and Xu, 2015. Caenagnathasia
sp. (Theropoda: Oviraptorosauria) from the Iren Dabasu Formation (Upper Cretaceous:
Campanian) of Erenhot, Nei Mongol, China. Vertebrata PalAsiatica. 53(4), 291-298.
Funston, Currie, Ryan and Dong, 2019. Birdlike growth and mixed-age
flocks in avimimids (Theropoda, Oviraptorosauria). Scientific Reports.
9:18816.
undescribed possible dromaeosaurine (Currie and Eberth, 1993)
Middle-Late Campanian, Late Cretaceous
Iren Dabasu Formation, Inner Mongolia, China
Material- teeth
Comments- Currie and Eberth (1993) state "Isolated dromaeosaurid teeth and bones are common in
the Iren Dabasu" and that "Most of these can be attributed to Velociraptor,
although some of the teeth suggest that there was a second, larger
species of an indeterminate dromaeosaurine dromaeosaurid." A pedal
phalanx II-1 (AMNH 6572) mentioned by Ostrom (1969) as being 20% larger
than Deinonychus may belong
to the same taxon, based on size.
References- Ostrom, 1969. Osteology of Deinonychus antirrhopus,
an unusual theropod from the Lower Cretaceous of Montana. Peabody Museum of
Natural History Bulletin. 30, 1-165.
Currie and Eberth, 1993. Palaeontology, sedimentology and palaeoecology of the
Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia, People s Republic
of China. Cretaceous Research. 14, 127-144.
undescribed Averostra (Chow and Rozhdestvensky, 1960)
Middle-Late Campanian, Late Cretaceous
Iren Dabasu Formation, Inner Mongolia, China
Material- (AMNH 6376) phalanx II-1 (AMNH online)
(AMNH 6556) metatarsal II (AMNH online)
(AMNH 6744) four caudal vertebrae, 8 distal pedal elements (AMNH online)
(AMNH 6756) metatarsal (AMNH online)
(AMNH 6757) limb fragments, metapodials, phalanx, fragments (AMNH online)
(AMNH 21552) femur
(AMNH 21565) elements
(AMNH 21588)
(AMNH 21774) fibula
(AMNH 21775) pedal phalanx ?II-1
(AMNH 21776) four proximal pedal phalanges
(AMNH 21780) four unguals
(AMNH 21782) manual ungual
(AMNH 21784) four caudal vertebrae
(AMNH 30245) two metatarsal II or IV shafts (AMNH online)
(AMNH 30247) posterior dorsal rib fragment (AMNH online)
(AMNH 30248) proximal anterior rib (AMNH online)
(AMNH 30249) partial coracoid (AMNH online)
(AMNH 30250) distal femur (AMNH online)
(AMNH 30251) proximal femur (AMNH online)
(AMNH 30252) distal femur (AMNH online)
(AMNH 30253) proximal femur (AMNH online)
(AMNH 30254) distal femur (AMNH online)
(AMNH 30255) astragalus (AMNH online)
(AMNH 30256) proximal tibia (AMNH online)
(AMNH 30257) proximal femur (AMNH online)
(AMNH 30258) distal tibia (AMNH online)
(AMNH 30259) proximal metatarsal (AMNH online)
(AMNH 30260) proximal metatarsal (AMNH online)
(AMNH 30298) acetabular fragment (AMNH online)
(AMNH 30299) proximal ischium (AMNH online)
(AMNH 30360) metatarsal III shaft (AMNH online)
(AMNH 80277) distal humerus (AMNH online)
(IVPP or PIN coll.) (small) three partial skeletons (Chow and Rozhdestvensky, 1960)
(IVPP and PIN coll.) <400 specimens (Currie and Eberth, 1993)
Comments- The AMNH specimens
listed here are from the online catalogue, which generally lacks
identification for ranks between order and family so that Theropoda
indet. material is listed as Saurischia. Yet none of the specimens are
likely to be sauropods given Gilmore (1933) never mentioned finding any
and to this day only a few elements have been reported (4 in the
Erenhot Dinosaur Museum coll., 7 from the Sino-Soviet expedition-
Currie and Eberth, 1993; Sonidosaurus).
Most would have been found during the April 22 to May 25 1923 Central
Asiatic Expedition, AMNH 6556 on April 30. The online catalog also
specifies AMNH 6756 was discovered in AMNH site 141. Many of these
specimens (AMNH 6556, 30245, 30247-30260, 30298-30299, 30360, 80277)
are listed as being from "8 mi. E. of station" which would place them
among Third Asiatic Expedition field sites 140-149, with AMNH 6757
listed as 9 miles east, so perhaps site 149. One exception is AMNH
6744, stated as being found at Elephant Camp (12 miles NW of the
station) by de Chardin, who was only on the 1930 expedition. Based on
the elements preserved, specimen numbers and locality of "8 mi. E. of
station", sevcral specimens (AMNH 30245, 30247-30260, 30298-30299,
30360) may belong to the two(+?) troodontid individuals noted in the
AMNH online catalog represented by specimen numbers AMNH 30261-30297,
30300-30318, 30320-30330 and 30336-30359.
Chow and Rozhdestvensky (1960) noted "three partially complete
skeletons of some small carnosaurian dinosaurs" discovered in the
June-July 1959 Sino-Soviet expedition, perhaps indicating
tyrannosauroids or dromaeosaurids. Currie and Eberth (1993) stated "A
rough tally of Sino-Soviet field identifications shows that ...
'theropods' (including large theropods, small theropods and segnosaurs,
but not ornithomimids) were more common (400 specimens)."
References- Gilmore, 1933. On the dinosaurian fauna of the Iren Dabasu
Formation. Bulletin American Museum of Natural History. 67, 23-78.
Chow and Rozhdestvensky, 1960. Exploration in Inner Mongolia - A
preliminary account of the 1959 field work of the Sino-Soviet
Plaeontological Expedition. Vertebrata PalAsiatica. 4(1), 1-10.
Dong, Currie and Russell, 1989. The 1988 field program of The Dinosaur Project. Vertebrata PalAsiatica. 27(3), 233-236.
Dong, 1992. Dinosaurian Faunas of China. China Ocean Press. 188 pp.
Currie and Eberth, 1993. Palaeontology, sedimentology and palaeoecology of the
Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia, People’s Republic
of China. Cretaceous Research. 14, 127-144.
Dong, 1993. The field activities of the Sino-Canadian Dinosaur Project
in China, 1987-1990. Canadian Journal of Earth Sciences. 30(10),
1997-2001.